Внутpеннee строение и образ жизни вымершего паразитического насекомого Saurophthirus Longipes PONOMARENKO, 1976 (INSECTA, APHANIPTERA, SAUROPHTHIROIDEA)
Введение
В работе дается характеристика дыхательной и
пищеварительной систем раннемелового насекомого Saurophthirus
longipes, описанного
А.Г.Пономаренко в 1976. Новизна работы заключается в том, что это первое
описание внутреннего строения не только вида S.longipes,
но и группы заврофтероидов в целом. Эти данные актуальны, так как позволяют
дать более точную трактовку образа жизни заврофтируса. Полученные результаты
могут быть полезны для решения таких фундаментальных вопросов, как эволюция
паразитизма, эволюция систем органов насекомых и становление связи
паразит-хозяин.
Авторы предыдущих публикаций сходятся во мнении
о том, что S.longipes
- это эктопаразит каких-то позвоночных с частично голой эластичной кожей,
скорее всего птерозавров. В данной работе эта гипотеза проверяется на примере
изучения анатомии древней блохи. Слабая склеротизация брюшка самок позволяет
довольно подробно охарактеризовать дыхательную и пищеварительную системы. В
ходе работы была выдвинута рабочая гипотеза о том, что для S.longipes
характерен гонотрофический цикл, что меняет представление о его биологии.
Цель работы: проверить гипотезу о
паразитировании Saurophthirus
longipes на птерозаврах,
изучив его внутренне строение.
Для достижения цели были поставлены следующие
задачи:
Дать характеристику пищеварительной системы Saurophthirus
longipes
Провести элементный анализ образцов по
содержимому кишечника
Дать характеристику дыхательной системы Saurophthirus
longipes
Проверить соответствие внутреннего строения и
предполагаемого ранее образа жизни
Основываясь на полученных данных, теоретически
обосновать образ жизни Saurophthirus
longipes.
Автор благодарна своему научному руководителю,
М.Е. Черняховскому (МПГУ) за свое знакомство с Палеонтологическим институтом
своему научному консультанту, заведующему лабораторией артропод ПИН РАН А.П.
Расницыну за курирование работы и определение жизненного пути, А.Г. Пономаренко
(ПИН РАН) за участие в обсуждении результатов работы, А.А. Прокину (ИБВВ РАН) и
Д.Е. Щербакову (ПИН РАН) за помощь в получении необходимой литературы, Р.А.
Ракитову (ПИН РАН) за прекрасные SEM
фотографии и проведенный элементный анализ, Е.А. Жегалло и Л.В. Зайцевой за
помощь в интерпретации результатов элементного анализа.
Работа поддержана Программой Президиума РАН № 22
«Эволюция органического мира и планетарные процессы».
Часть 1.Обзор литературы
.1 Характеристика ископаемых паразитов
Понятие хищник, паразит, паразитоид
В природе очень часто взаимоотношения между
организмами, когда один из них питается другим. Такое питание подразумевает три
варианта связи: хищник-жертва, паразитоид-хозяин и паразит - хозяин. Для того,
чтобы понять, с каким типом связи мы имеем дело, надо ответить на два вопроса:
умирает ли жертва (хозяин), и как быстро она (он) умирает. Среди насекомых
встречается все три типа взаимоотношений. Стрекозы, златоглазки, богомолы,
кузнечики, клопы - щитники, клопы-водомерки, жужелицы, божьи коровки и многие
другие нападают на жертву, тут же ее убивают и съедают. Оруссиды, тригоналиды,
всевозможные наездники и некоторые другие семейства перепончатокрылых и
веерокрылые относятся к паразитоидам. Они обитают в или на теле хозяина,
постепенно поедая его.
Это и приводит к смерти последнего. Паразиты, в
отличие от паразитоидов, не убивают своего хозяина, и их связь может быть еще
более длительной.
.2 Причины плохой сохраняемости ископаемых
паразитов
Ископаемые насекомые известны по копролитам с
силура, а по плохо определяемым обрывкам с девона. И хотя по молекулярным
данным эта группа должна быть еще более древней, образцы, которые можно с
уверенностью определить как насекомое, описаны только в карбоне2. Судить о том,
когда у насекомых появилась связь паразит-хозяин, довольно трудно. Паразиты
очень редко сохраняются в палеонтологической летописи. Во- первых, они часто
находятся внутри тела хозяина, из-за чего их нельзя опознать. Во-вторых,
довольно существенную часть жизненного цикла паразиты могут проводить вдалеке
от хозяина, и тогда сложно будет предугадать, является ли этот древний организм
паразитом. Тем более, по тофономическим причинам может сохраниться только
паразит или только хозяин (позвоночные и беспозвоночные, как правило,
сохраняются в разных условиях). В-третьих, для паразитов характерны мелкий
размер (особенно для вирусов или бактерий) и слабая склеротизация тела (для
червей), из-за чего их будет сложно опознать в ископаемом состоянии. Наконец,
эктопаразиты могут быть подвержены разложению вместе со скелетом хозяина, и,
поэтому, найдены не будут. Есть еще одно важное соображение: паразиты, как
правило, вымирают вместе со своим хозяином. Тогда, ископаемое паразитическое
насекомое должно иметь исчезнувшего хозяина. А если описывается связь
паразит-хозяин, где паразит вымер, а хозяин - нет,
скорее
всего, определение этой связи было ошибочным.
.3 Описание палеонтологических находок
факультативных паразитов
Ископаемые двукрылые
В палеонтологической летописи насекомых известны
и облигатные, и факультативные паразиты. К факультативным паразитам относятся
многие двукрылые, клопы, жуки, сетчатокрылые и скорпионницы. К облигатным -
вши, блохи и пухоеды. Далее будет представлено краткое описание
палеонтологических находок обеих групп.
Факультативные паразиты сидят на хозяине не
постоянно, а лишь иногда единовременно получают большую порцию пищи. Зачастую
такими паразитами являются только самки, а самцы питаются соками растений.
Самки тоже могут питаться растительными соками, и переходить на паразитический
образ жизни в период размножения. Так происходит у обыкновенных комаров. И
самка, и самец постоянно питаются сахарами растений, медовой росой или
экзофлорным нектаром. Но как только у самки
появляется
необходимость в питательных веществах для развития яиц, она переходит на
питание кровью млекопитающих и птиц. Получив достаточную порцию крови, она
откладывает яйца в стоячий водоем и снова переходит на питание нектаром.
В палеонтологической летописи двукрылые известны
с перми. И уже к триасу сформировались все современные группы. Появление
кровососов именно в это время связано с отсутствием на суши подходящей жертвы
ранее. Первые млекопитающие появились на территории восточной Европы (Thomasia,
Eoraetia
siegerty), Англии (Kuehneotherium,
Morganucodon) и Франции в
триасе. А оперенные динозавры и вовсе в верхней юре в
Китае(Aurornis
xui).
Москиты (Culicidae)
- это многочисленная группа комаров-кровососов почти всех современных
млекопитающих. Culicidae
представлены в летописи окаменелостей не менее чем 25 видами. Из них 11
известны в янтаре и 14 представлены окаменелыми остатками. По молекулярным
данным группа появилась в среднем триасе, а широко распространилась в течение
средней юры. На звание самого древнего представителя этой группы претендуют
Grauvogelia
аrzvilleriana из среднего триаса
и Burmaculex
antiquus из среднемелового
бирманского янтаря.
Кроме москитов описано более 250 видов Ceratopogonidae,
7 видов Corethrellidae3,
порядка 75 видов Psychodidae4,
80 видов слепней и бекасниц, из которых 28 - это лошадиные мушки6. Некоторые
другие группы, например Corethrellidae7,
тоже описаны, но не столь многочисленны.
Мухи семейства Simuliidae
описаны как одни из первых паразитов нелетающих динозавров и птиц, что
определили по строению их коготков. Представители другой группы двукрылых,
москитов (Phlebotominae)
найдены на шерсти млекопитающих из миоценового доминиканского янтаря8. Но все
это достаточно привычные сегодня паразиты. Среди двукрылых описаны и более
специфичные животные. Например, водная личинка мухи, предка современных Athericidae,
Qiyia jurassica,
паразитировавшая
в средней юре, Даухугоу, на саламандрах. У этой необычной личинки грудные
сегменты слиты, на их нижней стороне имеется грудная присоска, а от присоски
отходит 6 гребней с рядами щетинок, направленных вверх. Может быть, гребни -
это видоизмененные грудные конечности, которые увеличивают площадь всасывания.
На каждом сегменте брюшка имеются коренастые ложные ножки для закрепления на
теле хозяина с 4 крючками на концах. На заднем конце тела - две пары анальных
сосочков (современные мухи их используют для изымания кислорода из воды, играют
огромную роль в ионном обмене). Ротовой аппарат представлен колюще- сосущим
хоботком. В этих же отложениях найдено много ископаемых саламандр с тонкой
кожей за жабрами. На них и могла паразитировать Qiyia
jurassica.
Еще более удивительная группа- это Strashilidae,
представленная тремя видами: Strashila
incredibilis
Rasnitsyn, 19922, Strashila
daohugouensis
Huang et
al., 20133 и Vosila
sinensis
Vršanský
et Ren,
20104. Это бескрылые мезозойские насекомые, которые изначально были описаны в
отряде скорпионниц. Для них характерны длинные конечности, подобно современным
паразитам летучих мышей (мух Nycteribiidae
и Streblidae (Hippoboscoidea),
клопов Polyctenidae
(Cimicoidea) и клещей Spinturnicidae
(Gamasina)),
короткие 10-членниковые антенны, колюще-сосущий хоботок, доходящий до первого
брюшного сегмента, пятичленниковые лапки и слабо
склеротизированное
брюшко. Было высказано предположение, что это паразиты птерозавров. На
последнем сегменте брюшка имеются крючки. Они могли служить для закрепления на
жартве во время кровососания. Первичное описание основывалось только на
строении самцов. В дальнейшем в средней юре Китая обнаружили новые отпечатки,
среди которых были и самки с некоторыми особенностями строения, позволяющими
перенести этот вид в отряд двукрылых. Во-первых, у самок не оказалось таких же
специализированных для закрепления на хозяине задних конечностей. Т.е, это
всего-навсего средство удержания самки самцом при спаривании. Во-вторых, тело
страшилы в отличие от заврофтируса и тарвинии цилиндрическое, а не сплюснутое.
В-третьих, гениталии самца устроены скорее как у Chironomidae,
Sciaridae или некоторых Nymphomyiidae,
но не
как
у Mecoptera, ближайших
родственников блох.
Ископаемые сетчатокрылые
Вместе с двукрылыми в перми появились паразиты
из других отрядов насекомых. Например, Neuroptera.
Единственная паразитическая группа невроптеров - это Mantispidae.
Это облигатные паразиты пауков и перепончатокрылых. Личинки Mantispinae
паразитируют на яйцах паука, некоторые личинки паразитируют на самом пауке,
пока он еще не отложил яйца. На пауке личинка питается гемолимфой3. Всего
описано 14 видов мантиспид. Из них 5 описаны из янтаря: Micromantispa
cristata в меловом янтаре
Мьянмы, Doratomantispa
burmanica в бирманском
янтаре, Dicromantispa
electromexicana в
мексиканском янтаре, D.
moronei в доминиканском
янтаре, Feroseta
prisca в доминиканском
янтаре. Остальные
виды
найдены в месторождениях юрского, мелового периодов, эоцена и олигоцена. Два из
них (Mantispidiptera)
описаны Гримальди предположительно
Ископаемые жуки
Еще одна группа - жуки. Сами жуки появились в
карбоне или перми2. А вот паразитические формы, Meloidae,
паразиты прямокрылых и некоторых перепончатокрылых, по молекулярным данным
возникла в раннем мелу3. Всего описано три ископаемых личинки нарывников: Proplebeia
dominicana из Доминиканского
янтаря раннего миоцена, которая паразитировала на пчелах, сомнительная личинка
из Балтийского янтаря среднего эоцена и еще одна личинка из Балтийского янтаря.
Последняя находка - самая древняя. Сейчас нарывники на пчелах не паразитируют.
Так что эта связь между паразитом и хозяином была потеряна.
Среди других отрядов насекомых паразиты известны
у скорпионниц, клопов и некоторых других групп, которые здесь рассмотренны не
будут.
Описание облигатных ископаемых паразитов
Облигатные паразиты животных
Кроме того, что масса насекомых лишь временно
посещает своих хозяевах, можно вспомнить и немало облигатных паразитов.
Например, Phthiraptera.
Вши и пухоеды - эктопаразиты всех видов птиц и почти всех млекопитающих. В
настоящее время в этой группе описано более 5000 видов, а вот в ископаемом
состоянии их известно намного меньше. По молекулярным данным вши Anoplura
появились в конце мела1. На данный момент описано порядка 6 видов. Один из них,
Psittacobrosus
bechsteini, найден в гнездах
птиц и млекопитающих, оперении и меху. Некоторые другие находки имеют спорный
характер, и могут вскоре быть переописаны как клещи. Нахождение паразита на
хозяине среди насекомых известно крайне редко. Чуть ли не единственный пример -
это вошь Megamenopon
rasnitsyni, найденная в Германии.
Ее возраст 44 млн лет (эоцен). В передней части экземпляра сохранились остатки
крыла позвоночного, из чего ученые сделали вывод, что это паразит птиц.
Филогенетический анализ указал на то, что это перо предка современных
гусеобразных или куликов.
В Phthiraptera
описано одно удивительно ископаемое, которое, как и страшилиды, было описано
как паразит птерозавров. Это пухоед Saurodectes
vrsanskyi. Для него
характерны удлинные конечности, похожие на ноги страшилы (и современных
паразитов летучих мышей), но в отличие от последних завродектусы имеют
одночленниковую лапку. Мандибулы у
них
короткие и направлены вверх. По этим двум признакам насекомое отнесли к
пухоедам.
Столь же скудная на ископаемые находки группа
блох. В эоцене- миоцене описано 6 ископаемых блох: Palaeopsylla
klebsiana, Palaeopsylla
dissimilis, Peusianapsylla
baltica, Peusianapsylla
groehni из Балтийского
янтаря, Pulex
larimerius и Eospilopsyllus
kobberti из Доминиканской
республики. Все похожи на современных блох2
.Облигатные паразиты растений
Пару слов стоит сказать и об облигатных
паразитах растений. Бактерии, грибы, клещи, вирусы, нематоды и конечно
насекомые могут откладывать свои яйца в растения, индуцируя появление галлов,
мин или погрызов на коре. Это и есть паразитизм на растениях. Насекомое
защищает свое яйцо и обеспечивает питание личинке. Растению такие
взаимоотношения приносят вред, но не убивают его совсем. Галлы могут
образовывать цинипиды, галлицы, листоблошки, трипсы, разнокрылые стрекозы,
пилильщики, жуки, вши, листовертки и некоторые моли. Для определения
ископаемого паразита, который вызвал появление галла, сравнивают его форму,
размер, положение на листе, и, конечно, учитывают пищевые предпочтения самого
паразита. Галлы появлялись параллельно у разных групп насекомых. Первое
упоминание относится еще к каменноугольному периоду. Правда, такие галлы могли
образовать не только насекомые, но и клещи. Поэтому нельзя с уверенностью
сказать, что это первая находка галла насекомого. Широкое
распространение
галлов произошло только в позднем мелу, с расселением по планете
покрытосеменных.
Таким образом, ископаемые паразиты представлены
довольно большим количеством видов. Однако, большинство из них - факультативные
паразиты, в основном, двукрылые. Облигатные паразиты, такие как вши, пухоеды и
блохи, сохраняются намного хуже и поэтому в палеонтологической летописи
представлены буквально несколькими видами.
.4 Характеристика группы завроптероидов
Систематика группы
В столь немногочисленной группе ископаемых
облигатных паразитов существует целая группа удивительных насекомых,
систематическое положение которых до сих пор до конца не ясно. Это группа
мезозойских
«блох»,
где описан и Saurophthirus
longipes, которому
посвящена данная работа. Группа представлена тремя семействами: Saurophthiridae
Ponomarenko, 1986; Pseudopulicidae
Gao, Shih,
Ren, 2012 и Tarwiniidae
Huang, Engel,
Cai, Nel,
20132. Для представителей всех трех семейств характерны длинные прицепные ноги.
Именно по этому признаку их и объединяют. Ввиду их несомненного сходства и очевидно
также родства эти семейства были соединены в самостоятельное надсемейство
Saurophthiriodea
Ponomarenko, 19861. Это
позволяет обойтись без нечеткой формулировки мезозойские «блохи».
Описание первого представителя группы, Tarwinia
australis
История этой необычной группы блох началась еще
в 1970 году, когда Рик описал из нижнего мела (апта) Кунварры в Австралии
первого представителя T.
australis. В описании этого
насекомого автор усмотрел несколько удивительных черт: голова слабо сжата с
боков, антенны не укорочены, конечности необычно длинные, бедра не
прыгательное, а голень покрыта короткими щетинками. Рик предположил, что такие
конечности необходимы, чтобы цепляться за редкую шерсть какого-то животного2.
Описание S.
longipes
В 1976 году А.Г. Пономаренко описал найденное в
Байсе3 (неоком, вероятно, баррем Забайкалья) в единственном экземпляре
необычное бескрылое насекомое Saurophthirus
longipes
Ponomarenko, 1976 (рис. 1), в
котором, несмотря на серьезные отличия (крупное, не сжатое с боков тело,
длинные антенны, длинные, слабые, не прыгательные ноги, мягкое брюшко) усмотрел
сходство с блохами. Одной из причин такого определения была похожесть с
описанной ранее Tarwinia
australis
Riek,1970. Другое
важное соображение, на которое обратил внимание А.Г. Пономаренко - сочетание
колюще‐сосущего
хоботка и других свидетельств паразитического,кровососущего образа жизни
(мягкое раздувающееся брюшко и прицепные ноги) с гомономной грудью. Это
исключало родство с двукрылыми, основной альтернативой для кровососов среди насекомых
(поскольку родство со вшами и другими Paraneoptera
исключается пятичлениковыми лапками заврофтируса). Оставалось только сближение
со скорпионницами или с их потомками - блохами. Треугольная голова, длинные
ходящие конечности с двумя коготками на лапках, разделенный на две части
девятый сегмент брюшка действительно позволяют отнести заврофтируса к
скорпионницам. Но большинство авторов все же считают, что это блоха. А.Г.
Пономаренко отмечает сходство заврофтируса с различными паразитами летучих мышей
(Polyctenidae из
полужесткокрылых, Nycteribiidae
и Streblidae из двукрылых, Arixeniidae
из уховерток и Spinturnicidae
из клещей). Для всех этих паразитов характерны очень длинные, «циркульные»
прицепные ноги, которые позволяют им удерживаться на перепонке крыла при любых
его движениях. В мезозое группой, аналогичной летучим мышам были птерозавры. На
них и мог паразитировать заврофтирус.
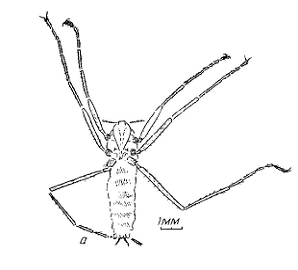


Рисунок 1. Внешнее строение
Saurophthirus longipes, голотип ПИН
№ 4210/1998: а - рисунок, б - фото (Пономаренко,
1976)
Особое семейство для Saurophthirus
было установлено только через десять лет в связи с описанием рода Saurophthiroides
Ponomarenko, 1986 из раннего
мела Западной Монголии, впоследствии оказавшегося представителем внешне
сходной, но далекой и тоже загадочной группы Chresmodidae
(Rasnitsyn, Quicke,
2002: 256, 366). Упомянутый там же (Rasnitsyn,
Quicke, 2002: 242) Saurophthirus
sp. из нижнемелового
местонахождения Семен в Забайкалье также основано на находке представителя Chresmodidae.
В этой же группе мезозойских «блох» в 1992 году
была описана Strashila
incredibilis, но позже она была
отнесена к отряду двукрылых3 (см.выше).
Таким образом, у началу XXI
века в группе мезозойсских «блох» было описано всего два насекомых: T.
аustralis и S.
longipes.
Все кардинально изменилось после описания трех
новых родов завроптироидовиз северовосточного Китая (пров. Внутренняя
Монголия). Это Pseudopulex
Gao et
al, 2012, Hadropsylla
Huang et
al., 2013 и Tyrannopsylla
Huang et
al., 20131. Семь
новых видов были описаны с промежутком буквально в пару лет. Из среднеюрских
отложений формации Цзюлоншань (Juilongshan
Fm) происходят P.
jurassicus
Gao, Shih
et Ren,
2012,
P. wangi
Huang, Engel,
Cal et
Nel, 2013 и Hadropsylla
sinica Huang,
Engel, Cai
et Nel,
2013 , а P. magnus
Gao, Shih
et Ren,
2012, P. tanlan
Gao, Shih,
Rasnitsyn
et Ren,
2014 и T. beipiaoensis
Huang, Engel,
Cal et
Nel, 2013 из
раннемеловых формации исянь (Yixian
Fm). Отсюда же описан
и Saurophthirus
exquisitus
Gao, Shih,
Rasnitsyn, et
Ren, 2013, второй вид
рода, ранее известного только из Забайкалья.
Сравнение всех завроптироидов
Из трех семейств заврофтироидов Pseudopulicidae
распространены в Китае и известны со средней юры до раннего мела, Saurophthiridae
найдены только в раннем мелу, но распространены шире - в Китае и Сибири
(Забайкалье), а для Tarwiniidae
существует единственная находка в раннем мелу Австралии. Saurophthiridae
отличаются от других семейств цилиндрическим (не вальковатым) телом самки,
сравнительно коротким
хоботком,
мягким брюшком самки с почти утраченной внешней сегментацией и длинными
полувтянутыми гениталиями самца. Соответственно Pseudopulicidae
характеризуются веретеновидным телом самки, мощным хоботком, более явственной
сегментацией брюшка самки и короткими, широкими не втяжными гениталиями самца. Tarwiniidae,
известные по единственному отпечатку самца, отличаются от обоих предыдущих
семейств слегка сжатым с боков телом, густой щеткой волосков на вершине и по
внешнему краю голеней и короткими втяжными гениталиями. Дополнительно тарвиния
характеризуется сравнительно слабо измененной головной капсулой и свободными
челюстными щупиками, а также мощными передними тазиками и голенями, тогда как у
двух других семейств голова шлемовидная (сужена к вершине, где расположены
маленькие глаза и где крепятся антенны), а щупики постоянно прилежат к хоботку.
Выборка в 9 видов позволяет проследить филогению заврофтироидов. Анализ
морфологии трех семейств, проведенный Гао с
соавторами,
позволил сделать вывод, что раньше всего в этой группе обособились тарвинии,
сохранившие относительно примитивное строение головной капсулы и щупиков, но
специализированных по строению ног и, возможно, гениталий. Из двух других
семейств псевдопулициды примитивны по менее дифференцированному опушению,
четкой сегментации брюшка у самки и наружным, не удлиненным гениталиям самца, и
более продвинуты по щупикам, более тесно ассоциированным со стилетами хоботка.
Напротив, заврофтириды специализированы по глубокой дифференциации опушения и
утрате ясной сегментации брюшка самки, по особенно тонким и длинным ногам и по
очень длинным, частично втяжным гениталиям самца.
Таким образом, за последние 5 лет представления
о завтофтироидах кардинально изменились. На данный момент описано девять видов
(Pseudopulex
jurassicus, P.
magnus, P.
tanlan, P.
wangi, Hadropsylla
sinica, Tyrannopsylla
beipiaoensis, Tarwinia
australis, Saurophthirus
longipes,
exquisitus),
относящихся к трем семействам (Saurophthiridae,
Pseudopulicidae и Tarwiniidae).
Из них три вида происходят из среднеюрской формации цзюлоншань в Китае (P.
jurassicus, P.
wangi и H.
sinica), четыре из
китайской нижнемеловой формации исянь (P.
tanlan, P.
magnus,
beipiaoensis
и S. exquisitus)
и по одному из нижнемеловых зазинской свиты Забайкалья (S.
longipes) и ископаемых
слоев Кунварры в Австралии (T.
australis). Среднеюрские
виды входят в состав так называемой биоты Яньляо (Yanliao),
известной пока только с территории СВ Китая, раннемеловые Китая и Сибири
представляют гораздо более широко распространенную биоту Жэхэ (Jehol),
обильно представленную и в Монголии1 . Положение австралийской Кунварры в этом
плане обособленное и пока точнее не идентифицированное.
Тенденции в эволюции надсемейства Saurophthiroidea
Общие тенденции в эволюции надсемейства Saurophthiroidea
выделить трудно, поскольку для каждого из трех его семейств характерны специализации
в разных направлениях. Наиболее специализированы несомненно заврофтириды:
псевдопулициды и тарвинии в разных отношениях более примитивны (см. выше). Если
же сопоставить надсемейство с современными блохами, эволюционная тенденция блох
в целом становится совершенно очевидной. Это уменьшение размеров тела, его
латеральная сплющивание и полная склеротизация брюшка, превращение прицепных
ног в прыгательные (их укорочение, расширение, усиление мускулатуры и
вооружения, сближение тазиков к средней линии тела), сильное укорочение антенн,
дальнейшая редукция глаз, развитие сложных систем ктенидиев для фиксации в
волосяном или перьевом покрове хозяев, уменьшение и интернализация мужских
гениталий.
Часть 2. Материалы и методы
.1 Краткое описание местонахождения Байса и её
биоты
Материалом для этой работы послужили отпечатки Saurophthirus
longipes, хранящиеся в
Лаборатории артропод Палеонтологического института им А.А.Борисяка РАН в Москве
и происходящие из нижнемеловых (возможно барремских) отложений зазинской свиты
местонахождения Байса в верховьях р. Витим Восточного Забайкалья (сборы ПИН РАН
1969, 1973, 1983 и 2000 гг.) Это отложения узкого и глубокого озера в межгорной
впадине Витимского вулканического плато, часто карбонатные (известняки,
мергели), местами битуминозные, а также
алевролиты,
песчаники, конгломераты1. В Байсе найдены остатки не менее
семейств
крылатых насекомых, относящихся к 25 отрядам. Многие из них автохтонны (50‐80
видов обитателей Байсинского озера) и часто более нигде не встречаются. Доминирующими
группами являются двукрылые,
поденки,
стрекозы, жуки и полужесткокрылые. Среди водных личинок особенно обильны комары
Chaoboridae («коретры»),
поденки Ephemeropsis
melanurus
Cockerell, стрекозы Hemeroscopus
baissicus
Pritykina и жуки Coptoclava
longipoda
Ping, а также домики
личинок ручейников. Среди других ископаемых встречаются перья птиц,
неопределимые кости, копролиты и следы тетрапод, остатки рыб, двустворчатых и
брюхоногих моллюсков, ракушковых рачков, конхострак, креветок, пауков, мшанок и
растений, окаменелая древесина, представляющие не только карбонатные
(ракушечные известняки и мергели) и битуминозные отложения, но и
песчаник,
брекчию, а также алевролиты и конгломераты.
.2 Описание использованного материала и
оборудования
Все 15 экземпляров S.
longipes, имеющиеся в
коллекции ПИН РАН, были изучены под микроскопом МПС-2 без спирта и под 95%
спиртом. Из них для работы были отобраны десять образцов: ПИН №№ 3064/1898
(голотип), 2378, 2379, 2380, 2381, 2383 и ПИН №№ 4210/6965, 6966, 2851,
. Для исследования только дыхательной системы
были использованы образцы под номерами: ПИН №№ 3064/2379, 2380, только
пищеварительной: ПИН №№ 4210/6966, 2851, 2852. Все исследуемые образцы
представлены отпечатками самок и были выбраны для изучения по причине слабой
склеротизации их брюшка, так как нормально склеротизованное брюшко самца не
позволяет увидеть строение его мягких тканей. Образцы были и отсняты под
бинокуляром Leica М165 С с
помощью камеры Leica
DFC 420. Соединение
фотографий проводилось в программах Helicone
Focus 5.1.28 и Kolor
Autopano
Giga 3.0.5.
Контрастирование фотографий проводилась в программах Adobe
Photoshop
CS6, PhotoScape
v3.6.2 и FastStone
Image Viewer
5.5. Фотографии высокого разрешения (SEM-
фотографии) получены при помощи сканирующего микроскопа TESCAH/Vega
XMU кабинета приборной
аналитики ПИН РАН (BSE
Detector). Для уточнения
гипотезы о гематофагии на четырех образцах под номерами: №№ 3064/2381, 2383,
1898 и № 4210/2851 был проведен элементный анализ поверхности. Результаты анализа
были занесены в карты. Рисунки по фотографиям сделаны в программе Corel
Draw X3.
При необходимости производилось дополнительное препарирование образцов с
помощью препаровальных игл, сделанных из заточенного круглого или трехгранного
надфиля.
Для работы были отобраны только те образцы,
которые представляли какой-либо интерес либо в плане изучения дыхательной
системы, либо пищеварительной, либо и той, и другой сразу. Поэтому десять
образцов считаются оптимальными для данных исследований. Использованные методы
позволили более точно интерпретировать строение дыхательной и пищеварительной
систем S.longipes.
Элементный анализ позволил выявить
спектр
веществ в кишечнике заврофтируса для уточнения ранее высказанной гипотезы о его
гематофагии.
Часть 3. Результаты
.1 Пищеварительная система
Характеристика пищеварительной системы S.
longipes (рис. 2) дается не
полно и только для тех ее отделов, которые удалось изучить на отпечатках.
Ротовой аппарат представлен колюще-сосущим хоботком с зазубренными стилетами,
как и у других заврофтироидов.
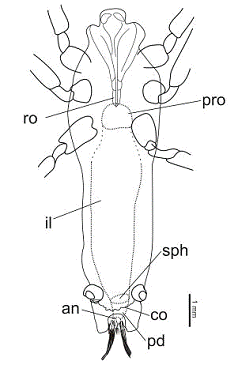
Рисунок 2. Пищеварительная система
Saurophthirus longipes
Условные обозначения здесь и далее: an
- анус, co -
воротничок, il - средний
кишечник, pd - задний
кишечник, pro -
провентрикулюс, ro
- хоботок, sph - сфинктер
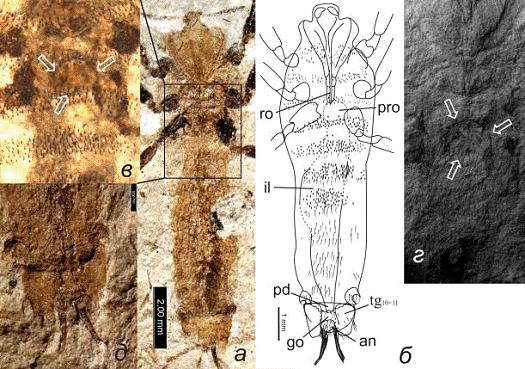
Рисунок 3. Saurophthirus
longipes, голотип ПИН №
3064/1898: а - фото, б - интерпретация пищеварительной системы, в - фото
провентрикулюса, г - SEM
фото провентрикулюса, д - фото задней части брюшка
Условные обозначения здесь и далее: полые
стрелки - провентрикулюс; go
- гонопор, tg10+11 -
слитые тергиты последних брюшных сегментов
Это один из признаков, указывающий на
паразитический образ жизни насекомого, так как зубцы могли служить для прокалывания
плотных покровов хозяина и заякоривания в его теле. Подробное описание ротового
аппарата давалось в более ранних публикациях1, поэтому в данной работе оно не
рассматривается.
Склеротизованные головная капсула, тергиты и
стерниты груди не позволяют видеть глотку и пищевод: единственное, что часто
бывает видно, это сильно склеротизированный провентрикулюс обычно на уровне
заднегруди (рис. 3в, г). Его ширина
составляет треть ширины грудного
тергита.
Никаких акант, располагающихся в полости провентрикулуса у блох и скорпионниц,
обнаружить не удается.

Рисунок 4. Saurophthirus
longipes, экз. ПИН №
3064/2378:а - фото, б - интерпретация пищеварительной системы, в - SEM
фото
Условные обозначения: полые указатели - средний кишечник,
белые стрелки - сфинктер
На некоторых образцах в пределах провентрикулюса
просматриваются более темные (иногда более светлые) образования неясной природы
(рис. 3в, г, 4в). Провентрикулюс очевидно не закреплен и, как и весь
кишечник,
может несколько изменять свое положение (смещаться вперед или в сторону; рис.
4б, в).

Рисунок 5. Saurophthirus
longipes, экз. ПИН №
3064/2381: а - фото, б - интерпретация пищеварительной системы, в - фото
провентрикулюса, г - фото задней части брюшка
От провентрикулюса отходит средний кишечник,
который резко расширяется и уже ко второму брюшному сегменту достигает 0,5
ширины брюшка (рис. 5, il).
Он, по-видимому, хорошо растяжим и может изменять свой диаметр в зависимости от
наполнения, которое предположительно жидкое и студенистое и зачастую собрано в
сгустки. Ни у одной из изученных самок кишечник не наполнен и брюшко не
растянуто, так что ширина кишечника почти на всем протяжении достигает 0,5 -
0,7 ширины
брюшка
(рис. 4, 5). Сужение кишечника происходит плавно к восьмому сегменту брюшка,
где диаметр достигает 0,3 ширины сегмента. На
всем протяжении кишечник прямой, без изгибов и выростов.
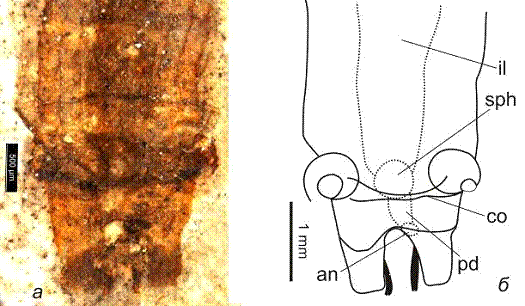
Рисунок 6. Saurophthirus
longipes, экз. ПИН №
4210/6965: а - фото, б - интерпретация пищеварительной системы
На восьмом сегменте брюшка средний кишечник
переходит в задний (рис. 3, 4, 5, 6 pd).
В некоторых случаях между средним и задним кишечником хорошо виден сфинктер
округлой формы размером с заднее дыхальце (рис. 6 - sph).
Видимого разделения заднего отдела кишечника на тонкую и прямую кишки нет.
Ректальный клапан и ректальные сосочки не просматриваются. Задний кишечник
открывается анальным отверстием вероятно на 10 сегменте брюшка, однако ни на
одном образце его расположение точно не было установлено. В одном случае (рис.
3б, д, go)
медиовентрально на 9 сегменте просматривается крупное отверстие, вероятно
женский гонопор.
Других элементов внутреннего строения S.
longipes (мальпигиевых
сосудов, слюнных желез, яйцеклада и др.) не обнаружено, вероятно, из-за их
слабой склеротизации.
.2 Элементный анализ
Элементный анализ образцов (рис. 7-8) показал в
кишечнике самок наличие углерода и серы, в отличие от окружающих частей тела и
породы вне ископаемых, где определяются кислород, кремний, алюминий, магний и
кальций (рис. 7а), а в некоторых случаях также калий и железо (рис. 7б).

Рисунок 7. Карта элементного анализа Saurophthirus
longipes: а - соотношение
разных элементов в средней части кишечника (голова справа), голотип ПИН №
3064/1898, б - соотношение углерода, калия и железа в средней части кишечника
(голова справа), экз. ПИН № 3064/2381
При этом сера сконцентрирована в задней части
кишечника, а в области провентрикулюса идентифицируются только ее следы (рис.
8а). Таким образом, предположение о наличии в кишечнике железа как индикатора
гемоглобина не подтвердилось. Чаще всего железо определяется равномерно
рассеянным по всей поверхности образца или, реже, везде вне кишечника заврофтируса
(рис. 8б).

Рисунок 8. Карта элементного анализа Saurophthirus
longipes: а - сравнение
соотношения углерода, серы и железа в средней части кишечника, экз. ПИН №
3064/2381, б - содержание углерода, серы и железа в провентрикулюсе, экз. ПИН №
3064/2381, в - то же, экз. ПИН №4210/2851
.3 Дыхательная система
На отобранных образцах было исследовано строение
дыхательной системы S.longipes
(рис. 9-13). Она представлена тремя парами хорошо развитых широких продольных трахейных
стволов, тесно сближенных в два блока общей шириной приблизительно 0,3 ширины
брюшка каждый.
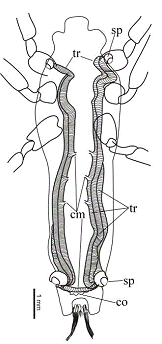
Риунок 9. Строение трахейной системы Saurophthirus
longipes

Рисунок 10. Saurophthirus
longipes, голотип ПИН №
4210/1998: a - фото, б
-интерпретация трахейной системы, в, г - SEM
фото передней и задней части тела
Условные обозначения здесь и далее, если не
сказано иначе: стрелки - передний тазик, сплошные треугольники - трахейные
стволы, полые треугольники - дыхыльце; cm
- комиссура, co -
воротничок, ro - хоботок, sp
- дыхальце, tr - трахейный
ствол
Трахейные стволы лежат дорзолатерально и
открываются на восьмом сегменте брюшка одной парой крупных дыхалец. Рострально
трахеи, сужаясь, прослеживаются до среднегрудных дыхалец, которые
идентифицированы предположительно: если идентификация корректна, то эти
дыхальца тоже велики. Во многих случаях
сохранились спиральные утолщения трахей (тенидии) (рис. 11).

Рисунок 11. Saurophthirus longipes,
экз. ПИН № 4210/6965: a - фото, б - интерпретация трахейной системы, в - SEM
фото участка двух трахейных стволов
Стволы в блоках не имеют видимых соединений (анастомозов),
лежат в основном параллельно, но могут изгибаться и переплетаться. Поэтому на
некоторых образцах просматривается лишь две пары трахей. В одном случае хорошо
видна широкая поперечная комиссура, соединяющая задние дыхальца и проходящая
дорзально по границе восьмого и девятого сегментов (рис. 12).

Рисунок 12. Saurophthirus
longipes, экз. ПИН №
3064/2381, а - фото, б - интерпретация трахейной системы, в - SEM
фото вершины брюшка с задней комиссурой
Условные обозначения: стрелки - анальный
гребень, остальные как на рис. 9
Другие трахеи, отходящие от продольных стволов
(возможно, поперечные комиссуры или висцеральные трахеи) видны редко (рис. 11,
13). Их расположение, по‐видимому,
метамерно: в известных случаях они отходят каудальнее сегментарных полос
шипиков, которые располагаются ровными поперечными рядами по спинной стороне
грудных и первых четырех брюшных сегментов. Трахейные стволы могут образовывать
петли и загибы около дыхалец (рис. 10, 12).
Рисунок 13. Saurophthirus
longipes, экз. ПИН №
3063/2378: а - фото, б - SEM
фото, в - интерпретация трахейной системы, г - SEM
фото головы и груди
На границе мягкого восьмого и склеротизованного
девятого тергитов просматривается толстая складка мембраны (рис. 10, 12).
Вероятно, она тоже связана с дыхательной системой и могла растягиваться при
отгибании девятого сегмента для освобождения крупных дыхалец при дыхании (см.
Обсуждение).
Часть 4. Обсуждение
.1 Пищеварителньая система S.longipes
Пищеварительная система насекомых
Пищеварительная система насекомых представлена
кишечником, состоящим из трех отделов: переднего, среднего и заднего. Передний
отдел сообщается с внешней средой ротовым отверстием, задний - анальным.
Передняя кишка делится на ротовую полость, глотку, пищевод, зоб и
провентрикулюс1. В ротовую полость открываются протоки слюнных желез. Глотка и
пищевод обеспечивают проглатывание пищи и ее прохождение в зоб (расширение или
слепое выпячивание пищевода) или прямо в среднюю кишку. Провентрикулюс
представляет собой сложную структуру, отделяющую пищевод от средней кишки.
Наличие хитиновой выстилки, отсутствующей в средней кишке, показывает
принадлежность провентрикулуса к переднему отделу кишечника. Провентрикулус
нередко снабжен мощной мускулатурой и толстой хитиновой выстилкой,
свидетельствующей о его участии в механической переработке пищи. В этих случаях
провентрикулус может быть различим у ископаемых даже на фоне склеротизованных
покровов. Внутренняя поверхность провентрикулюса может иметь вооружение для
дезинтеграции клеточного содержимого пищи2. Средний кишечник в отличие от
остальных отделов не имеет хитиновой интимы и поэтому хорошо растяжим. Он
выполняет функции выделения ферментов и всасывания продуктов пищеварения. Он
отделен от переднего отдела кардиальным клапаном, имеющим вид кольцевой
складки. Клапан препятствует проникновению пищи обратно в передний отдел
кишечника
или
регулирует этот процесс. В простейшем случае средняя кишка имеет вид прямой
трубки, не разделенной на вторичные отделы, но часто она удлинена
и
образует изгибы или, наоборот, укорочена и превращена в мешкообразный орган.
Задний кишечник подразделяется на три отдела:
пилорический отдел, тонкую и прямую кишку. Подобно передней кишке, он несет
хитиновую выстилку, но здесь она проницаема для воды и питательных веществ1.
Задняя кишка отделяется от средней пилорическим клапаном. За клапаном следует
сфинктер, который периодически открывается, чтобы пропустить содержимое средней
кишки. Между клапаном и сфинтером отходят мальпигиевы сосуды. Тонкая кишка
обычно длинная, извилистая, отделена от прямой ректальным клапаном. У
кровососов тонкий кишечник особенно длинный. Прямая кишка обычно короткая, в
норме несет ректальные сосочки.
Пищеварительная система у скорпионниц и блох
Строение кишечника у скорпионниц и блох,
ближайших родственников
S. longipes,
очень сходно. Провентрикулюс скорпионниц цилиндрический, темноокрашенный, несет
два ряда зубцов (акант) при входе и при выходе для размельчения частичек пищи.
Длина его зубцов в три раза больше толщины его стенки3 Аканты также
препятствуют обратному току полупереваренной массы во время испускания
скорпионницами пищеварительного сока в жертву для ее внекишечного
переваривания.У блох провентрикулюс также снабжен щетинками для разрушения
эритроцитов. Средний кишечник у блох и скорпионниц прямой и широкий. Задний
кишечник длинный и образует петли. Так же устроен задний кишечник у других
гематофагов, в частности у вшей.
Сравнение пищеварительной системы S.
longipes и блох
У S.
longipes задний кишечник
(проктодеум) очень короткий, прямой, относительно широкий и не разделен на
тонкую и прямую кишку; ректальные папиллы не просматириваются (рис. 3г). Такое
строение проктодеума для взрослых насекомых, особенно для Holometabola,
не характерно, тем более для гематофагов. Однако, по мнению авторов всех
предыдущих работ о заврофтироидах, последние были именно гематофагами. Это
противоречие может быть объяснено, по-видимому, только как педоморфоз, как
сохранение взрослым насекомым некоторых личиночных признаков, к которым, помимо
строения кишечника, может относиться также бескрылость и не склеротизованное
брюшко самки. Еще одним эффектом педоморфоза может быть отсутствие вооружения
провентрикулюса, столь характерное для блох и скорпионниц, ближайших
родственников S. longipes3.
Как известно, проктодеум в онтогенезе закладывается в виде простой трубки (для
блох см. Kessel E. L. The embryology of fleas
// Smifhsonian miscellaneous collections. 1939. Vol. 98(3). P. 1-90, fig. 7,
для скорпионниц Suzuki N. The comparative embryology of the Eumecoptera (Insecta,Mecoptera)
//PhD thesis at Tsukuba University. 1982. P.
1-239, fig. 194d).
Дополнительно гипотеза педоморфного упрощения кишечника заврофтируса основана
на данных о прямой, широкой и короткой, обычно лишенной ректальных сосочков
задней кишке многих личинок (у бабочек, пилильщиков), которая не всегда даже
дифференцирована на тонкую и прямую кишку (Richards
O.W.,
Davies R.G.
Imms’ general
textbook
of entomology.
Vol. 2: Classification and Biology // London:
Chapman and Hall, 1977. fig. 503; Omotoso O.T., Adedire C.O. Gross anatomy and
histology of the alimentary system of the larva of palm weevil, Rhynchophorus
Phoenicis Fabricius (Coleoptera: Curculionidae) // Journal of life sciences. 2010.
Vol. 4, N 1(26). P. 21-25.).
4.2 Элементный анализ
Сравнение результатов с полученными на других
гематофагах
Неожиданным для гипотезы гематофагии оказался и
результат проведенного нами элементного анализа содержимого кишечника S.
longipes. Анализ не показал
там повышенной концентрации железа как необходимого компонента крови
позвоночных. Это, казалось бы, вообще ставит под сомнение гипотезу о
гематофагии, хотя морфология заврофтироидов, как уже говорилось, достаточно
определенно указывает на их гематофагию. Однако ситуация не столь безнадежна.
Если сравнить наши данные с надежным примером идентификации гематофагии
насекомого на палеонтологическом
материале,
обнаруживается важное различие. В цитированной работе остатки кровососущих
комаров из среднеэоценовой формации Кишенен в Монтане, США продемонстрировали
не только ожидаемый явственный избыток железа, но и раздутое брюшко самки
комара, и мощный интенсивно окрашенный слой органики в брюшке, чего нет ни у
одного из наших заврофтирусов.
Объяснение полученных результатов элементного
анализа
Выйти из возникшего противоречия позволяют
следующие объяснения. Можно допустить, что в содержимом кишечника S.
longipes железа нет не
потому что он питался иной пищей, а потому что ни одна из исследованных самок
не была напитавшейся, что в содержимом их кишечника просто нет продуктов
переработки гемоглобина. Их либо уже нет (пища переварена, и ее остатки
выведены), либо еще нет (самки еще не питались). Остатки какого-то содержимого
тем не менее там присутствуют, как свидетельствует наличие избытка углерода и
серы, но они могут быть либо какими-то лишними, не подлежащими всасыванию
остатками, либо следами, например, какого-то дополнительного питания (как
известно, самки кровососущих комаров
охотно
пьют нектар), либо даже остатками пищи личинки. Тот факт, что сера
накапливается в задней части кишечника, может быть связан с тем, что всасывание
серосодержащих компонентов пищи у заврофтируса могло быть почему-либо
замедлено.
Тот факт, что исследуемый ископаемый материал
представлен только самками с пустым брюшком, т.е. еще не начинавшими или уже
отложившими яйца и потратившими все питательные везества, указывает на быстрое
протекание цикла, особенно его более рискованных этапов, когда самка с раздутым
брюшком особенно уязвима.
На данном этапе ни данные по строению
пищеварительной системы, ни данные полученные в ходе элементного анализа, не
противоречат ранее высказанной гипотезе о паразитировании заврофтируса на
птерозаврах на столько, чтобы ее как-то модифицировать.
Дыхательная система S.longipes
Дыхательная система насекомых
Как известно, дыхательная система насекомых
представлена трахеями, открывающимися в исходном варианте в 10 пар дыхалец,
расположенных по
бокам
средне‐
и заднегруди и восьми первых сегментов брюшка (голопневстическая трахейная
система). В типичном случае от каждого
дыхальца
отходит короткая трахея, которая дает начало трем другим поперечным, идущим
дорзально, вентрально и медиально. С функцией
соединения
поперечных трахей развиваются парные продольные стволы ‐
дорсальные,
латеральные и вентральные. В каждом сегменте от главных
стволов
отходят и ветвятся второстепенные трахеи, от которых далее отходят трахеолы,
поставляющие кислород непосредственно к клеткам. Часто количество дыхалец
меньше десяти, например, у многих личинок двукрылых они имеются лишь на
среднегруди и на восьмом сегменте брюшка (амфипневстическая) или только на
восьмом сегменте брюшка (метапневстическая трахейная система). Если трахейная
система вовсе не содержит открытых дыхалец, она называется замкнутой
(апневстической). Строение дыхалец изменчиво: они могут быть простыми
(отверстие с замыкательным аппаратом) и сложными (кроме обычного отверстия
имеется атриум, предохраняющий трахеи от заливания водой, попадания паразитов и
пыли).
Дыхательная система скорпионниц и блох
У блох (рис. 14) вдоль тела с каждой стороны
тянется по два продольных трахейных ствола, расставленных в груди и сближенных
в брюшке: судя по их положению в груди, это латеральный и вентральный стволы.
Трахейные стволы открывается в дыхальца на заднегруди и на восьми сегментах
брюшка и в каждом сегменте отсылают вверх и вниз поперечные дорзальные и
вентральные комиссуры. У скорпионниц, других предполагаемых современных
родственников Saurophthiroidea,
дыхательная система организована сходным образом и отличается в основном полным
набором
дыхалец (средне‐ и заднегрудные и
восемь пар брюшных)1.
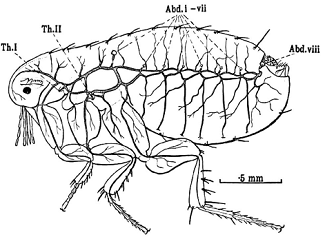
Рисунок 14. Трахейная система блохи Xenopsylla
(по Wigglesworth,
1935)
Условные обозначения: Th.1,
Th.2 - средне- и
заднегрудное дыхальца, Abd.i-vii,
Abd.viii
- брюшные дыхальца 1-го - 8-го сегментов
Сравнение дыхательной системы S.
longipes с блохами и
скорпионницами
Трахейная система S.
longipes оказалась гораздо
более специализированной, чем у современных скорпионниц и блох и, по крайней
мере, по числу дыхалец, чем у Pseudopulicidae
с их десятью парами крупных дыхалец1. У S.
longipes (и у других
заврофтирид, рис. 15) трахейная система, по-видимому, амфипнейстическая (если
не метапневстическая, см. выше) с очень крупной задней парой дыхалец, т.е. в
этом отношении она подобна таковой у многих личинок двукрылых.

Рисунок 15. Дыхальца и трахейные
стволы Saurophthirus exquisitus (Gao et al., 2013, fig. 2B, с изменениями)
Условные обозначения: Сплошные треугольники -
трахейные стволы, полые треугольники - дыхыльце
Важную специализацию представляет наличие с
каждой стороны тела по блоку из трех сильно расширенных трахейных стволов, что,
по‐видимому,
вообще не имеет аналогий у насекомых, хотя сами по себе три продольных
трахейных
ствола это примитивный признак. Тройной блок трахей мог
образоваться
путем смещения вентральных и латеральных продольных стволов вверх к дорзальному
стволу.
Известные аналоги строения дыхательной системы
Единственный известный функциональный аналог
описанной дыхательной системы это огромные продольные трахейные стволы у
метапневстических водных личинок жуков, например, у современных и вымерших
жуков плавунцов и их родичей (рис. 16).
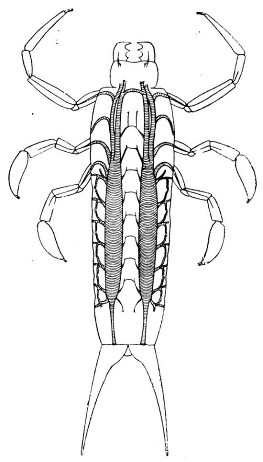
Рисунок 16. Дыхательная система Coptoclava
longipoda (Пономаренко,
1961)
У личинок плавунцов дыхательная система
представлена парой одиночных (не тройных) расширенных продольных стволов,
которые на первых личиночных стадиях открываются единственной парой дыхалец на
восьмом сегменте брюшка, и лишь на последних стадиях открывается
полный
набор дыхалец. Среди вымерших жуков Hydradephaga
можно выделить Coptoclava
longipoda
Ping, которую Пин Чжи
описал из формации лайян (Laiyang
Fm., СВ Китай),
аналога более известной китайской формации исянь и забайкальской зазинской
свиты. Позже этот вид был найден во многих приблизительно одновозрастных
отложениях Забайкалья, Монголии и Китая3. У личинок этого жука две толстые
трахеи (1/3 от ширины тергита каждая) тянутся вдоль всего тела и резко сужаются
при впадении в дыхальца на восьмом брюшном сегменте (рис. 15). Дыхальца
среднегруди и первых семи брюшных сегментов закрыты и на отпечатках имеют вид
зернышек. На
восьмом
сегменте стигмы увеличены в 2‐3 раза по
отношению к остальным.
Подобное строение дыхательной системы было
описано и для трех самых древних находок других личинок семейства Coptoclavidae5,
но в отличие от C. longipoda
у них просматривается среднегрудное дыхальце.
Предполагаемый для S.
longipes механизм дыхания
Важен механизм дыхания личинок плавунцов:
подплывая к поверхности воды, они отгибают вершину брюшка, открывая дыхальца.
При этом отогнутая вершина брюшка фиксируется на пленке поверхностного
натяжения, позволяя личинке свободно висеть в процессе дыхания, а в случае
опасности резко оторваться от нее и уплыть. Набрав кислород в трахеи,
личинка
разгибает брюшко, закрывая дыхальце, и уплывает. Описанная выше кожная складка
(воротничок) между восьмым и девятым тергитами брюшка
longipes,
позволяющая далеко отгибать вершину брюшка во время дыхания, углубляет
морфологическое сходство заврофтирид с водными личинками жуков и,
соответственно, сходство их предполагаемого способа дыхания.
.3 Изменение гипотезы об образе жизни S.
longipes
Причины модификации гипотезы об образе жизни
Описанные особенности трахейной системы Saurophthirus
заставляют пересмотреть существующие гипотезы об образе жизни этих насекомых.
Действительно, авторы работ по заврофтироидам сходятся во мнении о том,
что
это были эктопаразиты птерозавров либо каких‐то
других сравнительно
крупных
позвоночных с по крайней мере частично голой, эластичной кожей.
Об этом свидетельствуют такие признаки этих
насекомых, описанные в перечисленных публикациях, как:
сравнительно крупные размеры (порядка 1 см и
более),
мощный хоботок с зазубренными стилетами, слишком
толстый и грубый для прокалывания и высасывания насекомых или растений,
мягкое и сильно растягивающееся брюшко у самок и
нормально склеротизованное у самцов, что свидетельствует о единовременном
принятии самкой (но не самцом) большой порции жидкой пищи, необходимой для
развития
соответствующей порции яиц (как у иксодовых клещей и кровососущих комаров).
Известен даже случай сохранения такой самки с раздутым брюшком у псевдопулицид,
длинные и не слишком мощные прицепные («циркульные») ноги, позволяющие
насекомому продолжать питание, несмотря на значительную деформацию субстрата.
Такие ноги обычны у обитателей травостоя (особенно у комаров‐долгоножек),
но среди паразитов позвоночных они характерны лишь для кровососов на крыловой
перепонке летучих мышей.
В то же время амфи- или метапневстическая
дыхательная система с очень большими задними дыхальцами, открывающимися на
вершине брюшке (или перед вершиной, но тогда участок за дыхальцами способен
резко отгибаться, открывая дыхальца фактически на конце тела) свойственна
насекомым, обитающим в толще субстрата и имеющим лишь эпизодический доступ к
богатому источнику кислорода. А толстые продольные стволы, известные только у
водных личинок насекомых, еще более сужают круг возможных условий существования
заврофтируса. Это всё типичные водные адаптации, которые заставляет
предположить гораздо более тесную связь Saurophthiridae
с водой, чем это представлялось раньше. В свою очередь это делает понятным
пусть не частые, но довольно регулярные находки Saurophthirus
в захоронении, поскольку по тафономическим причинам (по закономерностям
захоронения: подробнее см. Жерихин В.В., Пономаренко А.Г., Расницын А.П.
Введение в палеоэнтомологию. М., 2008. Стр. 99-100.) в озерных отложениях,
откуда, если не считать янтарь, происходит огромное большинство ископаемых
насекомых, регулярно захораниваются только насекомые, живущие в воде или
летающие над водой.
Паразитирование на водном животном
Паразитизм на водном позвоночном мог бы
объяснить регулярное попадание Saurophthirus
в захоронение, но остатки подходящих хозяев
В средней юре Китая (Даохугоу) найдено водное
млекопитающее, аналогичное бобру или утконосу, но оно было покрыто густой
шерстью3 и тоже видимо не годилось в хозяева Saurophthirus.
Впрочем, и сам заврофтирус в юре не найден. Таким образом, связать паразита с
явными водными адаптациями с какими-либо водными хозяевами не удается.
Гонотрофический цикл
Выйти из возникшего противоречия позволяет то
обстоятельство, что разовый прием самкой эктопаразита большой порции крови,
необходимой для развития соответствующей порции яиц, предполагает существование
отработанного гонотрофического цикла. Такой цикл включает посещение
хозяина
для кровососания, затем уход с него в безопасное место для спокойного
переваривания и развития яиц, далее яйцекладка и, если повезет, повторение
цикла. Но если подобная циклика была свойственна заврофтироидам, если они вели
частично свободный образ жизни, можно допустить, что таким защищенным биотопом
для напитавшейся самки мог быть и водоем. Детализируя эту гипотезу, стоит
отметить, что для птерозавров общеизвестно широкое распространение рыбоядности
и, соответственно, тесная связь с водоемами, в том числе и с озерами (см.,
например, Zhou
Z.H.,
Wang Y.
Vertebrate
diversity
of the
Jehol Biota
as compared
with other
lagerstätten
// Science China
Earth Sciences.
2010. Vol. 53.
-1907.).
Поэтому для напитавшейся самки не представляло проблемы попадание в водоем,
равно как и обратное попадание на птерозавра, который в поисках добычи
курсирует низко над водой, задевая торчащие из нее стволики хвощей и других водных
растений. Личинки паразита могли бы быть постоянными обитателями водоема, а
самцы могли поджидать самок, когда те вылезали на растения. Предлагаемая
реконструкция, несомненно, носит экзотический характер, но по крайней мере она
явным образом не противоречит наблюдениям. К тому же увеличенные, хотя и не
олигомеризованные дыхальца самок Pseudopulicidae
также намекают на возможность водного образа жизни и этого более примитивного
семейства. Если предположение об активных среднегрудных дыхальцах самки заврофтируса
справедливо, эти дыхальца очевидно могли использоваться только в надводном
положении.
Альтернативная гипотеза Д.Е. Щербакова
В ответ на статью о дыхательной системе
заврофтируса1 сотрудник ПИН РАН Д.Е.Щербаков высказал свою альтернативную
гипотезу, которая предполагает постоянный эктопаразитизм заврофтируса и его
куклородность
(как
у мух надсемейства Hippoboscoidea)
и объясняет наличие у него гипертрофированных трахейных стволов связью с водой
их хозяев - птерозавров.
Эту гипотезу трудно принять по ряду причин.
Прежде всего, постоянные эктопаразиты даже связанных с водой амниот имеют
ничтожные шансы на захоронение в осадочных породах. Известно лишь два таких
примера, касающиеся пухоедов, несомненных или предполагаемых, и известных
каждый по одному экземпляру из той же Байсы и из среднего эоцена Германии3. Saurophthirus
longipes, найденный в Байсе
в числе 15 экземпляров, едва ли может быть поставлен в этот ряд.
С другой стороны, свойственная самкам
заврофтируса дыхательная система амфи- или метапнейстического типа с
гипертрофированными трахейными стволами характерна и объяснима лишь для условий
длительного пребывания под водой с редкими, но произвольными экскурсиями к
богатому источнику кислорода, подобно личинкам жуков- плавунцов. Постоянным
эктопаразитам эта редкая адаптация не нужна. Равным образом постоянным
эктопаразитам обычно свойственно и более или менее постоянное питание: в этих
условиях особенно трудно объяснить отсутствие следов крови в содержимом
кишечника всех четырех самок, проанализированных на этот предмет (см. выше).
Автор ссылается на слабость гипотезы
гонотрофического цикла дополнительно из-за обилия в ориктоценозе Байсы остатков
рыб рода Lycoptera
Müller,
1848, делавших байсинское озеро слишком опасным убежищем для заврофтируса. Дело
однако в том, что остатки ликоптеры обильны лишь в специфическом ориктоценозе
битуминозных сланцев
байсинского
разреза. В ориктоценозе мергелей опасным для заврофтируса хищником были скорее
вымершие плавунцы семейства Coptoclavidae,
а не рыбы.
Выводы
Ротовой аппарат S.longipes
представлен колюще-сосущим хоботком. Кишечник на всем протяжении прямой, что
объясняется явлением педоморфоза. На уровне заднегруди просматривается
сильносклеротизованный провентрикулюс без акант.
В результате элементного анализа показано
наличие в кишечнике углерода и серы и отсутствие железа, что объясняется тем,
что изученные самки не были напитавшимися. Ни данные по строению
пищеварительной системы, ни данные, полученные в ходе элементного анализа, не
противоречат ранее высказанной гипотезе о паразитировании S.longipes
на птерозаврах.
У изученного вида дыхательная система
представлена двумя блоками по три трахеи , которые тянутся по бокам тела и
открываются двумя парами дыхалец на среднегруди и восьмом сегменте брюшка.
Подобная дыхательная система характерна для водных личинок жуков плавунцов и их
древних родственников Captoclavidae.
Такое строение дыхательной системы мы объясняем
наличием у самки гонотрофического цикла
Наши данные позволили изменить ранее высказанную
гипотезу об образе жизни S.longipes.
Для древней блохи характерен гонотрофический цикл.
Список литературы
Алифанов
В.Р., Савельев С.В. Древнейший орнитомимозавр (Theropoda,
Dinosauria) с отпечатками
покровов из верхней юры России // Палеонтологический журнал. 2015. N
6. С. 71-85.
Беклемишев
В.Н. Биоценологические основы сравнительной паразитологии. М.: Наука, 1970.
Жерихин
В.В., Пономаренко А.Г., Расницын А.П. Введение в палеоэнтомологию. М.:
Товарищество научных изданий КМК, 2008.
Захваткин
Ю.А. Курс общей энтомологии. М.: Колос, 2001.
Пономаренко
А.Г. О систематическом положении Captoclava
longipoda
Ping (Insecta,
Coleoptera) //
Палеонтологический журнал. 1961. N
3. С. 67-72.
Пономаренко
А.Г. Coptoclava
(Coleoptera) - своеобразный
раннемеловой водный жук из Восточной Азии // Труды Совместной
Советско-Монгольской палеонтологической экспедиции 1975. Вып. 2. С. 122-139.
Пономаренко
А.Г. Новое насекомое из мела Забайкалья - возможный паразит летающих ящеров //
Палеонтологический журнал. 1976. N
3. С. 102-106.
Пономаренко
А.Г. Scarabaeiformes
incertae
sedis // Труды
совместной Советско-Монгольской палеонтологической экспедиции. 1986. Вып. 28.
Расницын
А.П., Стрельникова О.Д. Дыхательная система и биология нижнемелового
ископаемого Saurophthirus
longipes
Ponomarenko, 1976
(Insecta,
?Aphaniptera, Saurophthiroidea
stat. nov.)
// Палеонтологический журнал. 2017. N
2. С. 61-72.
Скобло
В.М., Лямина Н.А., Руднев А.Ф., Лузина И.В. Континентальный верхний мезозой
Прибайкалья и Забайкалья / под ред. Н.А.Логачева - Новосибирск: Издательство
Сибирского отделения Российской Академии Наук, 2001.
Шванвич
Б.Н. Курс общей энтомологии. Введение в изучение строения и функций тела
насекомых. М.; Л.: Советская наука, 1949.