Влияние молекулярного кислорода на спектрально-оптические свойства окрашенных растворов гемоглобина в пористой среде
Министерство
образования и науки Российской Федерации
Федеральное Государственное бюджетное
образовательное учреждение
высшего профессионального образования
"оренбургский государственный университет"
Физический
факультет
Кафедра
радиофизики и электроники
выпускная квалификационная работа
Влияние
молекулярного кислорода на спектрально-оптические свойства окрашенных растворов
гемоглобина в пористой среде
Оренбург 2015
ЗАДАНИЕ
на выполнение выпускной квалификационной работы
студентке Евминовой Светлане Николаевне
по направлению подготовки (специальности) 010707.65 - Медицинская физика
Тема выпускной квалификационной работы (Утверждена приказом № 1948 - С от
15.12.2014)
Влияние молекулярного кислорода на спектрально-оптические свойства окрашенных
растворов гемоглобина в пористой среде
Срок сдачи студентом ВКР "01" июня 2015 г.
Цель и задачи выпускной квалификационной работы
Целью данной работы является экспериментальное исследование влияния
молекулярного кислорода на спектрально-оптические свойства растворов
гемоглобина с применением разных красителей в кислородопроницаемых и пористых
средах.
Для выполнения цели работы необходимо решить следующие задачи:
.1 Изучить влияние молекулярного кислорода на спектрально-оптические
характеристики белковых молекул
.2 Изучить влияние параметров силикагеля на спектрально-оптические
свойства окрашенных раствором.
.3 Провести эксперименты по влиянию молекулярного кислорода на
спектрально-оптические свойства окрашенных растворов гемоглобина в пористой
среде.
Исходные данные к выпускной квалификационной работе
.1 Статьи и монографии по вопросам строения, функционирования и
особенностей молекулы гемоглобина, а также статьи о динамике процессов
присоединения молекулярного кислорода к молекулам гемоглобина;
.2 Приборы и оборудование: спектрофотометр Т-70, люминесцентная установка
на базе монохроматора МДР-204, лабораторные весы HR-60, лазер blue laser torch, кинетическая спектральная
установка.
.3 Программное обеспечение спектрофотометра Т-70, программное обеспечение
монохроматора ВДР-204, программный пакет MicroCAL Origin 7.5 Pro.
Перечень графического материала
Иллюстративный материал должен грамотно отражать все результаты дипломной
работы, включая выбор пористой среды, влияние молекулярного кислорода на
люминесценцию окрашенных растворов гемоглобина в силикагеле.
Дата выдачи и получения задания
Руководитель"17" декабря 2014 г. _________ В.Н.Степанов
Студент"17" декабря 2014 г. _________ С.Н.Евминова
Содержание
Введение
. Процесс взаимодействия гемоглобина с молекулами кислорода
1.1 Важность молекулярного кислорода в дыхательном процессе
.2 Роль гемоглобина как переносчика кислорода
.3 Пористая структура - силикагель
.4 Явление люминесценции
2. Описание экспериментальных установок и методик проведения
экспериментов
2.1 Люминесцентная установка с возможностью откачки воздуха
из кюветы с образцом
.1.1 Аппарат лазерный АТС 53-250
.1.2 Вакуумный мини-насос Милливак
.1.3 Монохроматор МДР-204
.2 Лазерный кинетический спектрохронограф с возможностью
откачки воздуха из кюветы с образцом
.2.1 Импульсный лазер LQ529
.2.2 Осциллограф цифровой GDS-840 С
.3 Спектрофотометр Т70
.4 Электронные лабораторные весы HR-60
.5 Приготовление растворов
.6 Методика заливки пленок
.7 Проведение экспериментов
3. Результаты экспериментов
3.1 Абсорбционный эксперимент
.2 Люминесцентный эксперимент с жидкими растворами красителей
и гемоглобина
.3 Статистический анализ люминесценции пористых образцов,
окрашенных раствором красителей и гемоглобина
.4 Люминесцентный эксперимент с пористыми образцами,
окрашенными раствором красителей и гемоглобина с откачкой воздуха
.5 Кинетический эксперимент с пленками силикагеля,
окрашенного растворами красителей с гемоглобином и покрытого ПВБ
Основные результаты и выводы
Заключение
Список использованных источников
Введение
Гемоглобин - белок эритроцитов, красных кровяных клеток, переносящий
молекулярный кислород от легких к тканям в организмах позвоночных животных.
Гемоглобин можно считать своего рода модельным белком, структура, свойства и
функции которого наиболее полно изучены по сравнению с другими белками на
протяжении последних 50 лет. Американский физик Хопфилд назвал его атомом
водорода современной биохимии, имея в виду, что изучение гемоглобина сыграло в
биохимии ту же роль, что и изучение атома водорода в физике. Гемоглобин
называют также почетным ферментом, поскольку исследования его структуры в
статике и динамике позволили значительно продвинуться в понимании механизмов
функционирования ферментов. Структура этого глобулярного белка известна в
деталях главным образом благодаря работам английского биофизика Макса Перуца,
который получил первые рентгеноструктурные данные еще в конце 40-х годов нашего
века. За эти исследования он был удостоен Нобелевской премии.
Большая часть знаний о физиологии человека и многие аспекты патологии
берут свое начало от исследований гемоглобина. Его белки изучались путем
кристаллографии структур, клонированием и секвенированием генов, изучением ДНК.
Стала известна роль гемоглобина в транспорте оксидов азота и комплексный контроль
альфа и бета глобиновых генов. Эти данные позволили начать использовать
гидроксимочевину для увеличения уровня фетального гемоглобина при
серповидно-клеточной анемии.
Изучение структуры белков методом рентгеноструктурного анализа началось в
конце 1930-х гг., когда Дж.Бернал, Д.Кроуфут-Ходжкин, Д.Филлипс и другие стали
получать первые рентгенограммы белковых кристаллов. В то время метод
рентгеноструктурного анализа применялся в основном для изучения структуры
простых молекул. Белковые молекулы были настолько сложными, что в течение
последующих 15 лет ученым так и не удалось добиться существенных успехов при
изучении их структуры. Однако сам метод за это время значительно
усовершенствовался с технической стороны.
Стремительный прогресс в области рентгеноструктурного анализа начался в
1953 г. с разработки Перуцем метода изоморфного замещения (введение атомов
тяжелых металлов в молекулы кристаллических белков). Этот прием позволил
использовать рентгеноструктурный анализ для исследования белка гемоглобина.
Пока Перуц разбирался с молекулой гемоглобина, Кендрю успешно применил
его метод тяжелых атомов для изучения структуры сравнительно простого белка
миоглобина кашалота.
В ранних работах по изучению структуры гемоглобина Перуц получал проекции
молекул на плоскости. Однако, как писал впоследствии Кендрю [1], анализ таких
изображений, являвшихся по существу лишь силуэтами очень сложных молекул, давал
мало сведений об их структуре. Было очевидно, что при дальнейшем использовании
метода необходимо ввести в него такие усовершенствования, которые бы позволяли
исследовать структуры в системе трех координат.
В результате работы с миоглобином при использовании метода изоморфного
замещения Кендрю в 1957 г. на основе рентгеноструктурного анализа впервые
получил трехмерную модель этой молекулы с разрешение до 6 Ȧ. Хотя такое разрешение было
недостаточным для выявления боковых цепей и даже конфигурации самой
полипептидной цепи, оно давало картину общего расположения полипептидной цепи и
группы гема, другими словами, позволяло определить в общем виде третичную
структуру молекулы. Как вспоминал позднее Кендрю, "мы увидели нечто, чего
никто ранее не увидел… Это была трехмерная структура молекулы белка во всей ее
сложности" [2].
Применив для обработки результатов анализа ЭВМ, в 1959 г. Кендрю
расшифровал пространственное строение молекулы миоглобина. Он построил также
модель, дающую представление о положении большинства из 2500 атомов этой
молекулы.
Между тем, продолжая свои исследования, Перуц с сотрудниками получил
также трехмерное изображение молекулы гемоглобина лошади с разрешением до 5,5 Ȧ. Расшифровка структуры этой более
крупной белковой молекулы была значительно облегчена результатами, полученными
при изучении третичной структуры миоглобина кашалота. Оказалось, что структура
миоглобина чрезвычайно близка к третичной структуре каждой из четырех
субъединиц гемоглобина. Вот как об этом вспоминал сам Перуц: "Летом 1959
года, почти через 22 года после того, как я получил первую рентгенограмму
гемоглобина, мы смогли наконец построить трехмерную карту электронной плотности
гемоглобина с разрешающей способностью 5,5 ангстрем, подобно той, которая на
два года раньше была получена для миоглобина. И как только цифры со счетчика
были перенесены на контурную карту, мы убедились в том, что каждая из четырех
цепей гемоглобина по форме очень напоминает единственную цель миоглобина.
Бета-цепь и миоглобин были вообще идентичны, а альфа-цепь отличалась от них
лишь более коротким поперечником одной маленькой петли. Миоглобин был
экстрагирован Кендрю из мышц кашалота. Гемоглобин был получен из крови лошадей.
Позднейшие исследований показали, что миоглобин тюленя и лошади, гемоглобин
человека и коровы имеют также сходное строение" [3].
Молекулы гемоглобина человека состоят из плотно упакованных протеинов,
состоящих из симметричных спаренных димерных полипептидных цепей: альфа и бета-
глобинов, в тетрамерных структурно-функциональных единицах. Альфа2/бета2
молекулярные формы - основные структурные единицы гемоглобина взрослого
человека. Основная функция гемоглобина у млекопитающих - транспорт кислорода от
легких к тканям; он вовлечен в специфический транспорт трех других газов:
углекислого газа, монооксида углерода (угарного газа) и нитрида азота (NO).
Функциональное значение гемоглобина главным образом определяется
особенностью крепления аминокислотных цепей в глобиновых белках, включающих 7
витков в альфа цепи и 8 витков в бета цепи.
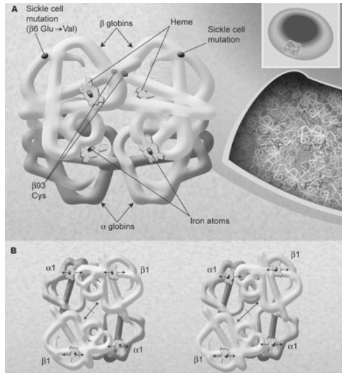
Рисунок 1 - Рентгеноструктурный анализ молекулы гемоглобина, показывающий
ее высокую компактность в эритроците
На рисунке 1 под буквой А изображено положение β-спирали (показано в виде трубок) в
каждой β-единице - одна слева и одна
повернута на 180°, справа - показано, как 4 гемовых группы с атомами железа
связывают молекулы газа. Также показано место серповидной мутации в мутантной β-глобиновой цепи и цистеин в 93
положении. Молекулы гемоглобина в эритроцитах, как показано на вставке справа,
плотно упакованы (концентрация 34 г/дЛ) и мало контактируют с растворителем.
Это позволяет кислороду не только эффективно перемещаться в каждой клетке, но и
влиять на химический состав молекулы. Например, полимеризации гемоглобина в
серповидной клетке при незначительной деоксигенации, на рисунке под буквой В
показаны изменения четвертичной структуры гемоглобина в тетрамере при переходе
от окси конформации (слева) к дезокси конформации (справа). Атомы железа
смещены относительно плоскости групп гемма и центральная полость между β-цепями, открываясь, облегчает связи
с 2.3 BPG.
Гемоглобин - один из наиболее хорошо изученных белков. Десятки лет
исследований гемоглобина во многих лабораториях мира привели к значительному
прогрессу в описании и понимании физических, химических и биологических
аспектов его функционирования. Работы Макса Перуца и его сотрудников в
Кавендишской лаборатории (Кембридж, Великобритания) внесли огромный вклад в
исследование этого белка. Однако важность этих работ касается не только
гемоглобина. Они послужили основой развития современных представлений о
механизмах ферментативного катализа, связав непосредственно кинетику и
термодинамику биохимических реакций с динамикой конформационных изменений
макромолекул белка. Если отвлечься от непосредственной практической пользы
полученных результатов для медицины, фармакологии, то фундаментальное значение
работ по изучению механизма функционирования гемоглобина заключается в
стимулировании прогресса в установлении законов протекания важнейших процессов:
ферментативного катализа и внутриклеточной трансформации энергии в
биологических системах.
Так как одной из основных функций гемоглобина является перенос молекул
кислорода, то наибольший интерес представляет исследование взаимодействия
гемоглобина с молекулярным кислородом. Исследования в данной области проводятся
традиционно оптико-спектроскопическими методами. В ряду крупных научных центров
проводятся изыскания в данном направлении, что указывает на актуальность данной
дипломной работы. Специфика данной дипломной работы заключается в том, что все
происходящие эффекты наблюдаются в специальных пористых средах. Некоторые
органы человека и других животных можно рассматривать как пористые среды, в
связи с чем произведена аналогия между исследуемыми образцами и некоторыми
органами человека. В качестве аналога пористой среды используется среда - силикагель.
В рамках дипломной работы сделан литературный обзор на тему исследования,
проведены серии оптико-спектроскопических исследований, получены данные
экспериментов и сделан ряд выводов.
1. Процесс
взаимодействия гемоглобина с молекулами кислорода
1.1 Важность
молекулярного кислорода в дыхательном процессе
Дыхание - это окислительно-восстановительный процесс, идущий с
образованием АТФ, при котором роль доноров водорода (электронов) играют
органические или неорганические соединения, а акцепторами водорода (электронов)
всегда служат неорганические соединения. Если конечным акцептором электронов
является молекулярный кислород, то такой дыхательный процесс называется
аэробным дыханием.
Очевидно преимущество кислородного (аэробного) типа энергетики перед анаэробиозом.
Количество энергии, выделяющейся при окислении данного питательного вещества
кислородом, в несколько раз превышает энергию, выделяющуюся при его окислении,
например, пировиноградной кислотой, используемой в качестве окислителя при
таком распространенном типе брожения, как гликолиз. Такие соотношения
обусловлены различиями в окислительно-восстановительных потенциалах пары
"кислород/вода" (+ 0,82 В) и "пировиноградная кислота/молочная
кислота" (- 0,19 В). Если учесть, что потенциалы основных субстратов
дыхания и брожения больше или равны - 0,7 В, то максимальные разности
потенциалов окислителя и восстановителя для дыхания будут составлять 0,7 В +
0,82 В = 1,52 В и для гликолиза 0,7 В - 0,19 В = 0,51 В. [4]
История открытия кислорода, как и азота, связана с продолжавшимся
несколько веков изучением атмосферного воздуха. О том, что воздух по своей
природе не однороден, а включает части, одна из которых поддерживает горение и
дыхание, а другая - нет, знали еще в 8 веке китайский алхимик Мао Хоа, а позднее
в Европе - Леонардо да Винчи. В 1665 английский естествоиспытатель Р. Гук
писал, что воздух состоит из газа, содержащегося в селитре, а также из
неактивного газа, составляющего большую часть воздуха. О том, что воздух
содержит элемент, поддерживающий жизнь, в 18 веке было известно многим химикам.
Шведский аптекарь и химик Карл Шееле начал изучать состав воздуха в 1768. В
течение трех лет он разлагал нагреванием селитры (KNO3, NaNO3) и другие
вещества и получал "огненный воздух", поддерживающий дыхание и горение.
Но результаты своих опытов Шееле обнародовал только в 1777 году в книге
"Химический трактат о воздухе и огне". В 1774 английский священник и
натуралист Дж. Пристли нагреванием "жженой ртути" (оксида ртути HgO)
получил газ, поддерживающий горение. Будучи в Париже, Пристли, не знавший, что
полученный им газ входит в состав воздуха, сообщил о своем открытии А. Лавуазье
и другим ученым. К этому времени был открыт и азот. В 1775 Лавуазье пришел к
выводу, что обычный воздух состоит из двух газов - газа, необходимого для
дыхания и поддерживающего горение, и газа "противоположного
характера" - азота. Лавуазье назвал поддерживающий горение газ oxygene -
"образующий кислоты" (от греч. oxys - кислый и gennao - рождаю;
отсюда и русское название "кислород"), так как он тогда считал, что
все кислоты содержат кислород. Давно уже известно, что кислоты бывают как
кислородсодержащими, так и бескислородными, но название, данное элементу
Лавуазье, осталось неизменным. На протяжении почти полутора веков 1/16 часть
массы атома кислорода служила единицей сравнения масс различных атомов между
собой и использовалась при численной характеристике масс атомов различных
элементов (так называемая кислородная шкала атомных масс).
Молекулярный кислород (газообразный или жидкий) - парамагнитное вещество,
в каждой молекуле О2 имеется по 2 неспаренных электрона. Этот факт
можно объяснить тем, что в молекуле на каждой из двух разрыхляющих орбиталей
находится по одному неспаренному электрону. Энергия диссоциации молекулы О2
на атомы довольно высока и составляет 493,57 кДж/моль.
В свободном виде встречается в виде двух модификаций О2
("обычный" кислород) и О3(озон). О2 - газ без
цвета и запаха. При нормальных условиях плотность газа кислорода 1,42897 кг/м3.
Температура кипения жидкого кислорода (жидкость имеет голубой цвет) равна
-182,9°C. При температурах от -218,7°C до -229,4°C существует твердый кислород
с кубической решеткой (-модификация), при температурах от -229,4°C до -249,3°C
- -модификация с гексагональной решеткой и при температурах ниже -249,3°C -
кубическая -модификация. При повышенном давлении и низких температурах получены
и другие модификации твердого кислорода.
При 20°С растворимость газа О2: 3,1 мл на 100 мл воды, 22 мл
на 100 мл этанола, 23,1 мл на 100 мл ацетона. Существуют органические
фторсодержащие жидкости (например, перфторбутилтетрагидрофуран), в которых
растворимость кислорода значительно более высокая.
Высокая прочность химической связи между атомами в молекуле О2
приводит к тому, что при комнатной температуре газообразный кислород химически
малоактивен. В природе он медленно вступает в превращения при процессах
гниения. Кроме того, кислород при комнатной температуре способен реагировать с
гемоглобином крови (точнее с железом II гемма), что обеспечивает перенос кислорода от органов дыхания к другим
органам.
Реакции биологического окисления, более эффективные, чем древние
энергетические процессы брожения и гликолиза, снабжают живые организмы большей
частью необходимой им энергии. Исключение составляют облигатные анаэробы,
например, некоторые паразиты, для которых кислород является ядом. Использование
кислорода, обладающего высоким окислительно-восстановительным потенциалом, в
качестве конечного акцептора электронов в цепи дыхательных ферментов, привело к
возникновению биохимического механизма дыхания современного типа. Этот механизм
и обеспечивает энергией аэробные организмы [5].
Наличие на Земле кислорода является одним из факторов возможности
существования жизни на нашей планете.
Кислород в составе воздуха через дыхательные пути попадает в легкие.
Концы самых мелких бронхов в легких заканчиваются множеством тонкостенных
легочных пузырьков альвеол - это 500 миллионов пузырьков диаметром 0,2 мм.
Здесь и происходит газообмен. Кислород из легочных пузырьков проникает в кровь,
а углекислый газ из крови - в легочные пузырьки.
Таким образом, венозная кровь обогащается кислородом и превращается в
артериальную. Кислород связывается с гемоглобином, который содержится в
эритроцитах, насыщенная кислородом кровь поступает в сердце и выталкивается в
большой круг кровообращения. По нему кровь разносит кислород по всем тканям
организма.
Поступление кислорода в ткани обеспечивает их оптимальное
функционирование, при недостаточном же поступлении наблюдается процесс
кислородного голодания - гипоксии [6].
Переносимый кислород находится в крови в двух состояниях. Небольшая часть
кислорода находится в растворенном состоянии в плазме. Коэффициент
растворимости кислорода в крови невелик и составляет (при температуре 37°С и
парциальном давлении кислорода 100 мм ртутного столба) 0,3 об.%. Это значит,
что каждые 100 мл нормальной крови могут переносить в растворенном состоянии
лишь 0,3 мл кислорода, что явно недостаточно для поддержания жизнедеятельности
организма человека.
В процессе эволюции был выработан принципиально другой механизм переноса
кислорода кровью. Этот механизм связан с наличием в крови специального сложного
белка (хромопротеида), способного обратимо присоединять молекулярный кислород.
В организме человека и высших животных таким хромопротеидом является
гемоглобин, содержащийся в эритроцитах.
Гемоглобин присоединяет кислород в среде с высоким парциальным давлением
кислорода и отдает кислород в среду с низким парциальным давлением. Каждый
грамм гемоглобина в нормальных условиях может присоединять 1,34 мл кислорода.
Поскольку нормальное содержание гемоглобина в крови составляет 145-170 г/л, то
100 мл крови могут переносить в связанном с гемоглобином состоянии 19-21 мл
кислорода [7].
В данной работе проводилось, в частности, воздействие лазерным излучением
на пористую среду (с гемоглобином) с высаженными на нем молекулами красителя
(люминесцентной метки).
В работе [8] исследована кинетика замедленной флуоресценции, вызванной
аннигиляцией триплетных возбужденных центров с мигрирующими 1Δg-возбуждениями молекулярного
кислорода.
Нелинейное тушение возбужденных электронных триплетных (Т) состояний
молекул красителей, внедренных в кислородосодержащие матрицы, возникает в
результате реакции аннигиляции Т- и 1Δg(О2)-возбуждений [9].
Временнáя зависимость концентрации Т-центров nT(t|n0) для различных начальных плотностей n0 этих частиц различна, и это легко обнаруживается при
изменении накачки системы. Экспериментально измеряемые величины, являющиеся
функциями или функционалами от nT, сохраняют чувствительность к вариациям n0. В работах [10] и [11] подчеркивалась нетипичная форма
временнóй зависимости интенсивности
замедленной флуоресценции (ЗФ) IDF(t) ряда
красителей, сопровождающей Т - 1Δg-слияние (кумуляцию энергии). Кроме
того, было установлено, что горбообразный характер кинетической кривой свечения
связан с сенсибилизированным заселением 1Δg - состояний в реакции
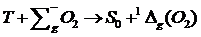 .(1.1)
.(1.1)
В начальный момент времени концентрация 1Δg -возбуждений nΔ равна нулю, а сигнал ЗФ нарастает в
течение некоторого времени tm. В [12] показано, что временные законы nT(t) и nΔ(t) формируются как суперпозиции локальных кинетик, получивших
специфическое развитие в пространственных областях с различной плотностью n0. В то же время в [13], установлена возможность
мгновенного (10 нс << τt,
τΔ,
где τt,
τΔ -
времена жизни Т-состояния красителя и 1Δg-состояния молекул О2
соответственно) однородного измерения концентрации nT с помощью дополнительного лазерного
импульса в произвольный момент времени t0 в результате лазероиндуцированной необратимой цепочки
переходов Т1 → Тm ↝Sn.
1.2 Роль
гемоглобина как переносчика кислорода
Гемоглобин - железосодержащий дыхательный пигмент крови позвоночных и
многих беспозвоночных животных, осуществляющий перенос кислорода от органов
дыхания к тканям организма. В крови позвоночных и некоторых беспозвоночных
гемоглобин содержится внутри эритроцитов в растворенном состоянии.
Молекула гемоглобина позвоночных животных состоит из белка - глобина и
железосодержащей группы - гема. В состав гема входят четыре протопорфириновых
кольца, каждое из которых содержит атом двухвалентного железа. Молекулярный вес
гемоглобина - 66 000- 68 000. Физиологическая функция гемоглобина как
переносчика кислорода основана на его способности обратимо связывать кислород в
зависимости от концентрации последнего в крови. В присутствии кислорода железо
гема связывает одну молекулу кислорода, при этом гемоглобин превращается в
оксигемоглобин. При взаимодействии гемоглобина с окисью углерода (например, при
отравлении этим газом) образуется более стабильный комплекс -
карбоксигемоглобин.
Продуктами распада гемоглобина являются многочисленные железопорфириновые
комплексы. При этом происходит полное отделение гема от белка (хромопротеида);
это отделение протекает с превращением железа в трехвалентную форму. Получаемый
железопротопорфирин называется гемином, а его соединения - геминодериватами.
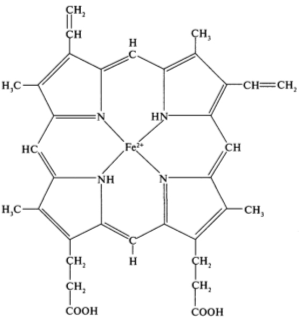
Рисунок 2 - Структура гема гемоглобина
Молекула гемоглобина состоит из четырех субъединиц: двух α
и двух β
- и соответственно
содержит четыре полипептидные цепочки двух сортов. Каждая α-цепочка содержит 141, а β-цепочка - 146 аминокислотных
остатков. Таким образом, вся молекула гемоглобина включает 574 аминокислоты.
Хотя аминокислотные последовательности α- и β-цепочек различны, они имеют
практически одинаковые третичные пространственные структуры. Собственно говоря,
приведенные выше детали структуры относятся не к гемоглобину, а к его белковой
компоненте - глобину. Каждая субъединица гемоглобина содержит одну небелковую
(так называемую простетическую) группу - гем. Гем представляет собой комплекс Fe(II) с протопорфирином. Структура гемма представлена на рисунке
2.
Группировка гема представляет собой сложную копланарную циклическую
систему, состоящую из центрального атома, который образует координационные
связи с четырьмя остатками пиррола, соединенными метановыми мостиками (= СН -).
В гемоглобине железо обычно находится в состоянии окисления (2+).
Четыре субъединицы - две α и две β - соединяются в единую структуру
таким образом, что α -субъединицы контактируют только с β -субъединицами и наоборот, как
показано на рисунке 3.
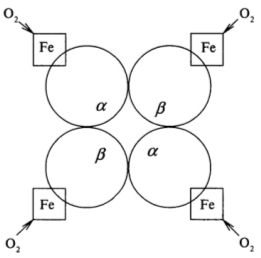
Рисунок 3 - Схематичное изображение четвертичной структуры гемоглобина:
Fe - гем гемоглобина
Как видно из рисунка 3, одна молекула гемоглобина способна переносить 4
молекулы кислорода. И связывание, и освобождение кислорода сопровождается
конформационными изменениями структуры α - и β -субъединиц гемоглобина и их
взаимного расположения в эпимолекуле. Этот факт свидетельствует о том, что
четвертичная структура белка не является абсолютно жесткой.
Атом железа может образовать шесть координационных связей. Четыре связи
направлены к атомам азота пиррольных колец, оставшиеся две связи -
перпендикулярно к плоскости порфиринового кольца по обе его стороны. Гемы
расположены вблизи поверхности белковой глобулы в специальных карманах,
образованных складками полипептидных цепочек глобина. Гемоглобин при нормальном
функционировании может находиться в одной из трех форм: феррогемоглобин (обычно
называемый дезоксигемоглобином или просто гемоглобином), оксигемоглобин и
ферригемоглобин (называемый также метгемоглобином). В феррогемоглобине железо
находится в закисной форме Fe(II), одна из двух связей,
перпендикулярных к плоскости порфиринового кольца, направлена к атому азота
гистидинового остатка, а вторая связь свободна. Кроме этого гистидинового
остатка, называемого проксимальным (соседним), по другую сторону порфиринового
кольца и на большем расстоянии от него находится другой гистидиновый остаток -
дистальный гистидин, не связанный непосредственно с атомом железа.
Взаимодействие молекулярного кислорода со свободным гемом приводит к
необратимому окислению атома железа гемма [Fe(II) → Fe(III); гем → гемин]. В дезоксигемоглобине глобин
предохраняет железо от окисления.
Обратимое присоединение кислорода (оксигенация), позволяющее гемоглобину
выполнять свою основную функцию переносчика, обеспечивается возможностью
образовать прочные пятую и шестую координационные связи и перенести электрон на
кислород не от железа (то есть окислить Fe2+), а от имидазольного кольца проксимального гистидина.
Для связывания кислорода с гемоглобином характерна кооперативность: после
присоединения первой молекулы кислорода связывание последующих облегчается. В
этом проявляется так называемый аллостерический эффект [15].
Стоит заметить, что лазерное излучение оказывает влияние на процесс
оксигенации гемоглобина. Авторы работы [16] измеряли величину насыщения
артериальной крови кислородом первой фаланги пальца с помощью
высокочувствительного быстродействующего пульсоксиметра. Третью фалангу пальца
подвергали облучению He-Ne лазера (20 мВт).
На рисунке 4 представлено изменение величины насыщения артериальной крови
кислородом при воздействии лазерного излучением.
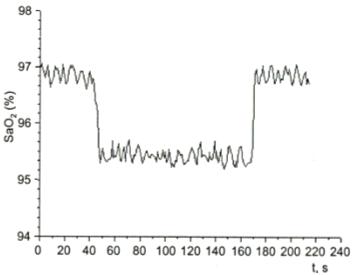
Рисунок 4 - Влияние лазерного излучения на насыщение артериальной крови
кислородом
На кривой насыщения регистрируются изменения от дыхательных циклов: 40
секунд - начало воздействия, 170 секунд - окончание. Снижение насыщения
гемоглобина синхронное с воздействием, демонстрирует дополнительное
освобождение кислорода в результате фотодиссоциации гемоглобина. Это не может
быть связано с улучшением микроциркуляции. Все происходит слишком быстро и
синхронно. Таким образом, это значит, что низкоинтенсивное лазерное облучение
высвобождает кислород в месте облучения. Авторы говорят о
лазерно-индуцированной оксигенации тканей. Речь идет о селективном повышении
локальной концентрации кислорода в тканях. Авторы объясняют этот эффект сдвигом
кривой диссоциации оксигемоглобина. И обосновывают это совпадением спектра
поглощения гемоглобина и оксигемоглобина с длиной волны He-Ne лазера. То есть, по мнению авторов, оксигемоглобин является
акцептором фотона.
Большой интерес для исследователей представляют особенности поведения
молекул газов (лигандов) в гемовом кармане гемоглобина и миоглобина. В работе
[16] рассмотрены механизмы диффузии лигандов в миоглобине, строение которого
очень сходно со строением β-субъединицы молекулы гемоглобина.
Результат расчетов Д. Кейза и М. Карплюса в 1979 году оказался по тем
временам несколько неожиданным. Выяснилось, что скорость диффузии лиганда в
белке чрезвычайно чувствительна к виду межатомных потенциалов взаимодействия,
определяющих конформационную подвижность. Конформационная подвижность
обусловлена возможностью вращения молекулярных групп вокруг одинарных
С-С-связей [17]. В вакууме при повороте на полный угол преодолеваются обычно
три потенциальных барьера высотой ~ 2-4 ккал/моль. В плотноупакованной белковой
глобуле эти вращения сильно заторможены из-за стерических препятствий, и,
казалось бы, ими можно пренебречь и рассматривать только небольшие колебания
атомов около локальных положений равновесия. Расчет динамики связывания лиганда
с миоглобином показал, что в этом случае энергия активации диффузии составляет
~ 100т ккал/моль, что примерно в 10 раз больше экспериментальной величины, и
процесс при комнатных температурах практически заморожен. Этот результат
полностью опроверг представление о белковой глобуле как об апериодическом
микрокристаллике. Оказалось, что включение в расчет конформационных степеней
свободы совершенно принципиально для белков и снижает энергию активации
диффузии лиганда до приемлемого значения ~ 10 ккал/моль. Более того, оказалось,
что в структуре миоглобина можно выделить два канала для проникновения лиганда
из раствора в гемовый карман. Эти каналы работают с вероятностью около 60 и 40%
соответственно и по своей структуре представляют систему флуктуационно
открывающихся щелей (дверей), образованных аминокислотными остатками. На
рисунке 5 схематически показан путь миграции СО в молекуле гемоглобина из
гемового кармана параллельно гемовой плоскости через участок альфа-спирали Е по
наиболее эффективному каналу. Молекула СО сначала преодолевает участок между
гистидином HisE7 и валином ValEll, а затем между HisE7 и треонином ThrElo с дальнейшим выходом в растворитель.
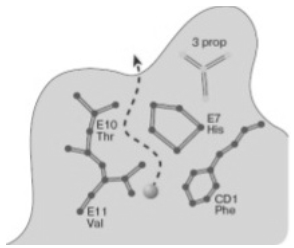
Рисунок 5 - Схематическое изображение одного из каналов для диффузии
лиганда в миоглобине
На рисунке 5 шариком указано место расположения связанного СО в гемовом
кармане. Пунктир показывает путь выхода лиганда в растворитель. Плоскость
рисунка параллельна гемовой плоскости и находится над ней на расстоянии 3,2 Ȧ. Обозначены положения аминокислотных
остатков с указанием спирального участка и номера остатка в спирали (HisE7, ValE11, ThrE10);
положение первого аминокислотного остатка в неспирализованном участке,
соединяющем спирали С и D (CD1Phe); участок одной из пропионатных боковых цепей,
присоединенных к молекуле гемма (3 prop).
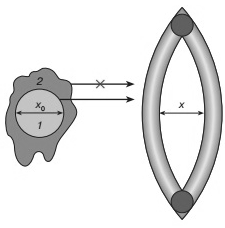
Рисунок 6 - Модель диффузии в структурированной среде (белке)
При раскрытии флуктуационной щели на величину х>x0 субстрат 1 может диффундировать сквозь щель. Диффузия
субстрата 2 сильно затруднена, так как требуется большая деформация щели
(большая энергия активации).
Рисунок 6 иллюстрирует общую схему процесса диффузии через флуктуирующую
щель, образованную элементами структуры белковой глобулы. Суть состоит в том,
что для акта диффузии необходимо, чтобы просвет во флуктуирующей щели был не
меньше, чем ванн-дер-ваальсов размер лиганда. Энергия активации диффузии при
этом будет определяться соответствующей энергией раскрытия щели. В жидкостях на
форму флуктуационной полости или дырки практически не накладывается существенных
ограничений. В белковой глобуле это не так. Здесь имеется относительно жесткий
упругий каркас, образованный спиральными элементами структуры. Подвижные
боковые группы обеспечивают формирование дырки для диффузионного проникновения
лиганда. Но форма этой дырки или щели диктуется локальной геометрией белковой
структуры. В твердых телах геометрические ограничения еще более жесткие, но в
этом случае в отличие от белка нет подвижных боковых групп, обеспечивающих
раскрытие щелей с приемлемой энергией активации. Таким образом, белковая
глобула представляет собой структурированную среду, в которой подвижная часть
похожа на вязкую жидкость, но возможные формы флуктуационных полостей и щелей
ограничены упругим каркасом [19]. Проникновение внутрь белковой глобулы молекул
(лигандов), геометрия которых не соответствует этим формам, будет сильно
затруднено, так как потребует более широкого раскрытия щелей и соответственно
значительно большей энергии активации. На этом этапе с точки зрения физики и
закладываются основы стереоспецифичности биохимических процессов.
1.3 Пористая
структура - силикагель
Силикагель - высушенный гель кремневой кислоты пористого строения с
сильно развитой внутренней поверхностью. Эта особенность обуславливает
ценнейшие свойства силикагеля - адсорбента, носителя каталитически активного
вещества и катализатора.
Получается при подкислении растворов силикатов щелочных металлов с
последующей промывкой и высушиванием образовавшегося геля:
Na2SiO3 + 2HCl →
2NaCl + H2SiO3↓,
H2SiO3 → SiO2 + H2O.
Существуют несколько способов получения силикагеля. Силикагель получают
взаимодействием силиката натрия с кислотой, щелочного силиката с солью аммония
из концентрированных золей коллоидного кремнезема; разбавлением растворов при
низких значениях pH и температурах
ниже комнатной; гидролизом соединений кремния (хлорид кремния, ортокремниевые
эфиры).
Силикагель имеет огромную площадь поверхности (800 м2/1 г),
состоящую из групп - SiOH,
расположенных на расстоянии 0,5 нм друг от друга. Эти группы являются активными
центрами, причем активность конкретной партии силикагеля зависит от числа и
активности таких центров. В активном адсорбенте, то есть таком, из которого
адалена адсорбированная на его поверхности вода, многие центры будут активны.
Такая активация происходит при нагревании геля до 150-2000С.
При нагревании до более высокой температуры в интервале 200-400°С
активность теряется в результате образования связей Si-O, происходящего с
отщеплением воды. Эта стадия, однако, обратима. При нагревании выше 400°С
размер поверхности силикагеля необратимо уменьшается.
Активные центры взаимодействуют с полярными растворенными веществами
главным образом за счет образования водородных связей. [20]
В зависимости от гранулометрического состава, формы частиц и характера
пористости силикагели обозначают четырьмя буквами: первая буква характеризует
размер гранул, вторая- (всегда С)- силикагель, третья- размер пор, последняя
форму частиц. Так, крупный силикагель мелкопористый гранулированный обозначают
КСМГ, мелкий силикагель мелкопористый кусковой - МСМК. Средние фракции
силикагеля называют "шихтой" и обозначают: ШСМК, ШСКГ, ШСМГ.
Индикаторный силикагель (ГОСТ 8984-58) -силикагель, пропитанный солями
кобальта. В зависимости от влажности среды он изменяет цвет от светло-голубого
до розового.
Выпускается силикагель в виде шариков или кусочков неправильной формы с
зернами размером в пределах 0,1-7,0 мм. В зависимости от аппаратурного
оформления рекомендуют следующий гранулометрический состав силикагеля: 0,1-0,25
мм - для процессов с кипящим слоем адсорбентов; 0,5-2,0 мм - для жидкофазных
процессов и процессов с движущимся слоем адсорбента; 2,0 - 7,0 мм - для
процессов в газовой фазе со стационарным слоем адсорбента.
Косвенной характеристикой структуры является насыпная плотность. Силикагель
мелкопористый имеет плотность около 700 г/л, а крупнопористый - от 400 до 500
г/л. Механическая прочность как от истирания так и от раздавливания также
различна: у мелкопористого она более высокая.
Промышленный силикагель содержит некоторое количество примесей - оксидов
алюминия, железа, кальция и других металлов. Некоторые из них активные
катализаторы и они способствуют протеканию крекинга при регенерации; в
результате на поверхности силикагеля образуется кокс, снижающий активность
поглотителя. Учитывая это, для осушки газов, в которых присутствуют
высококипящие углеводороды, используют более чистые силикагели с наибольшим
содержанием диоксида кремния.
Динамическая адсорбционная способность силикагеля по влаге зависит: 1 -
от размера зерна; 2 - скорости потока; 3 - содержания влаги в газе. [21]
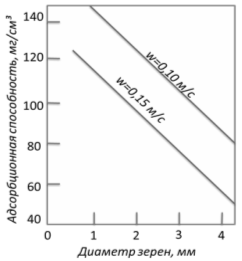
Рисунок 6 - Динамическая адсорбционная способность слоя мелкопористого
силикагеля (h=0,3 м) при разном размере зерна и
скорости потока (W, м/с). Осушка до
точки росы -60°С
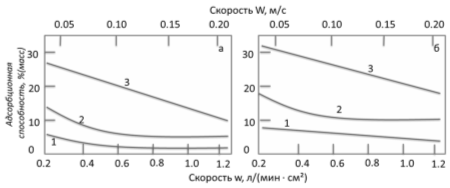
- крупнопористый силикагель; 2 - среднепористый; 3 - мелкопористый
Рисунок 7 - Динамическая адсорбционная способность слоя силикагелей (h=1 м) при разной скорости потока: а)
осушка для "проскоковой" концентрации, соответствующей точки росы tр=-40°С; б) то же, t=0°С
Ввиду гидрофильных свойств поверхности силикагеля его часто используют
для осушки воздуха, углекислого газа, водорода, кислорода, азота, хлора и
других промышленных газов.
Способность силикагеля поглощать значительные количества воды существенна
также для осушки различных жидкостей, в особенности в том случае, когда
обезвоживаемая жидкость плохо растворяем воду. Силикагели служат также
осушителями при консервации оборудования для предохранения его от коррозии.
Наряду с водой силикагель хорошо сорбирует пары многих органических
веществ. Этим его свойством пользуются для улавливания паров ценных
органических растворителей - бензина, бензола, эфира, ацетона и т.п. из
воздуха, бензола из газовых коксовых печей и бензина из природных газов.
С помощью геля кремневой кислоты проводится хроматографическое разделение
и анализ смесей, что основано на избирательности адсорбционного действия
силикагеля по отношению к веществам различной химической природы. Приведенные
примеры, далеко не охватывающие всех областей практического применения
силикагеля, характеризуют лишь главные направления, на которых особенно четко
проявляются его свойства. Интерес к силикагелю связан с сочетанием в нем ряда
ценных качеств: высокой адсорбционной способности, избирательности
адсорбционного действия, способности подвергаться многократной регенерации без
потери адсорбционной активности, относительно большой прочности зерен,
термостойкости, возможности получения его в гранулированном и порошкообразном
состоянии и др.
Важнейшим преимуществом силикагеля по сравнению с природными пористыми
материалами (пемза, асбест) является возможность изменения его структуры в
процессе формирования. Это обстоятельство является особенно важным потому, что
степень и характер пористости силикагеля обусловливают эффективность его
применения в различных процессах. При данной пористой структуре адсорбционная
активность определяется концентрацией адсорбируемого вещества и размером его
молекул.
Для хроматографического разделения веществ выбор величины удельной
поверхности и размеров пор зависит от свойств разделяемых молекул: для низших
углеводородов нужна большая поверхность и более узкие поры, для высших - низкая
поверхность и широкие поры. Хорошее разделение неполярных газов, которые
адсорбируются в основном благодаря неспецифическим дитерсионным
взаимодействиям, достигается в случае тонкопористых силикагелей со средним
диаметром пор не более 20 Ȧ. Для разделения легких углеводородов
пригодны силикагели диаметром пор от 50 до 200 Ȧ. Силикагели, у которых средний
размер пор больше 500 Ȧ, можно использовать для
газохроматографического разделения жидких смесей, в частности углеводородов.
Макропористые силикагели с низкой удельной поверхностью могут найти широкое
применение как носители неподвижных жидких и твердых фаз в газовой
хроматографии, в катализе, при адсорбции высокомолекулярных соединений и
полимеров из растворов. [22]
Пористая структура силикагеля, как и других гелеобразных адсорбентов,
формируется также и в процессе сушки. Объем геля при сушке уменьшается в десять
и больше раз. Поскольку пористая структура жестких силикагелей при смене
элементов не меняется, то не наблюдаются и изменения селективности
разделительной системы, если только геометрия растворенных полимерных молекул
не зависит от вида растворителя. Пути регулировки пористой структуры силикагеля
позволяют изменить удельные поверхности при одновременном изменении пористости
или получать адсорбенты с одинаковой удельной поверхностью, но разными объемом
и радиусом пор. Таким образом, Планк и Дрейк пришли к заключению, что пористая
структура силикагеля определяется размерами и плотностью упаковки составляющих
гель частиц. [23]
1.4 Явление люминесценции
Люминесценция - излучение, представляющее собой избыток над тепловым
излучением тела и продолжающееся в течение времени, значительно превышающего
период световых колебаний.
В отличие от рассеяния света, при люминесценции между поглощением и
испусканием происходят промежуточные процессы, длительность которых больше периода
световой волны. Однако критерий сравнения длительности этих процессов с
периодом световой волны недостаточен, чтобы, например, отделить резонансное
рассеяние от так называемой резонансной флуоресценции. При большом времени
жизни возбуждённого состояния акт резонансного рассеяния длится долее периода
световых колебаний, как и процессов когерентного испускания света, системой
атомов. Однако в этих процессах сохраняются определенные соотношения между
фазами поглощённой и испущенной световых волн, в то время как при люминесценции
эта корреляция утрачивается. Поэтому целесообразно отделять люминесценцию от
других процессов по времени фазовой релаксации поляризации среды.
Существует большое количество разнообразных как органических, так и
неорганических веществ, обладающих люминесцентной способностью, свойства
которой могут быть существенно неодинаковыми. Эти различия вызывают
необходимость в классификации явлений люминесценции.
В основу первой попытки такой классификации была положена
продолжительность процесса излучения [24]. В результате такой классификации все
известные виды люминесценции были разделены на два больших класса.
По длительности свечения различают флуоресценцию (быстрозатухающую
люминесценцию) и фосфоресценцию (длительную люминесценцию). Это деление
условное, оно зависит от временного разрешения регистрирующих приборов. Иногда
термины "флуоресценция" и "фосфоресценция" используют,
чтобы отличить люминесценцию, происходящую при переходах с синглетных уровней,
от переходов, происходящих с метастабильных триплетных уровней. [25]
По механизму элементарных процессов различают резонансную, спонтанную,
метастабильную, или вынужденную, и рекомбинационную люминесценцию.
В основу второго типа классификации были положены различные виды
возбуждения люминесценции: фотолюминесценция [26], атомная флуоресценция [27],
катодолюминесценция [28], рентгенолюминесценция [29], радиолюминесценция,
электролюминесценция [30], хемилюминесценция.
В основу третьего вида классификации, предложенного С.И.Вавиловым,
положена кинетика самого процесса люминесценции [31]. Согласно ему свечение
разделяют на резонансное, спонтанное, вынужденное и рекомбинационное.
Люминесцировать могут вещества во всех агрегатных состояниях - газы и
пары, растворы органических веществ, стёкла, кристаллические вещества; основное
условие - наличие дискретного спектра. Вещества с непрерывным энергетическим
спектром (например, металлы в конденсированном состоянии) не люминесцируют, так
как в них энергия возбуждения непрерывным образом переходит в теплоту. Кроме
того, для возникновения люминесценции вероятность излучательных переходов
должна превышать вероятность безызлучательного. Соотношение между этими
вероятностями определяет эффективность люминесценции. Интенсивность
люминесценции зависит от интенсивности возбуждения, поэтому не может служить
характеристикой люминесценции. Более однозначная характеристика - выход
люминесценции - отношение энергии люминесценции к поглощённой энергии
возбуждения (при фотолюминесценции - квантовый выход люминесценции - отношение
числа испущенных и поглощённых квантов света). [32]
В данной работе в качестве люминесцентной метки были использованы
органические красители эритрозин, эозин G и родамин 6G. Выбор этих красителей
не случаен и обусловлен высокой интенсивностью люминесцентного сигнала, а также
традиционностью использования этих красителей в биологических исследованиях.
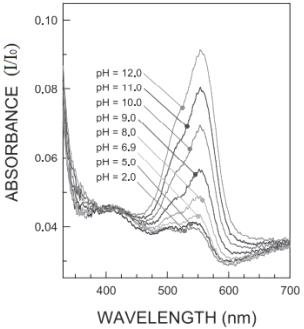
Рисунок 8 - Зависимость спектра поглощения водного раствора красителя
эритрозина от pH растворителя
Зависимость на рисунке 8 получена в работе [33]. Как видно из рисунка,
пик поглощения красителя эритрозина наблюдается на длине волны 525 нм.
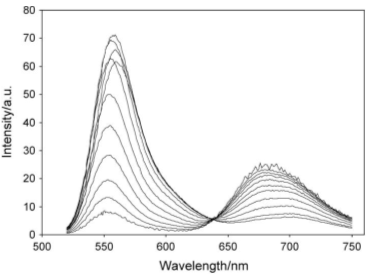
Рисунок 9 - Спектр замедленной флуоресценции эритрозина как функция
температуры
На рисунке 9 представлены спектры замедленной флуоресценции красителя
эритрозина в пленках сахарозы при температурах 5, 15, 25, 35, 45, 55, 65, 75,
95 и 100°С. Располагаются графики по порядку сверху вниз в окрестности длины
волны 670 нм. Эта зависимость получена в работе [34]. Максимум спектра быстрой
флуоресценции красителя эритрозина наблюдается на длине волны 553 нм.
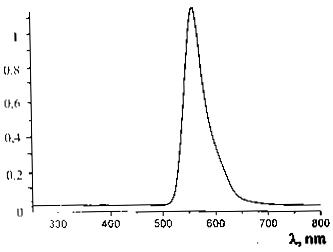
Рисунок 10 - Спектр люминесценции чистого водного раствора родамина 6G при возбуждении лазером диодной накачки
с λ=532
нм
Максимум люминесценции родамина 6G приходится на 552 нм.
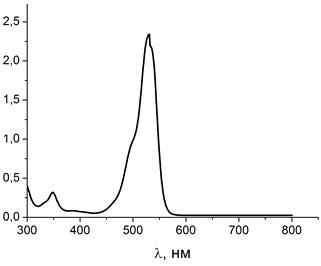
Рисунок 11 - Спектр поглощения чистого водного раствора родамина 6G
Максимум поглощения родамина 6G приходится на 530 нм.
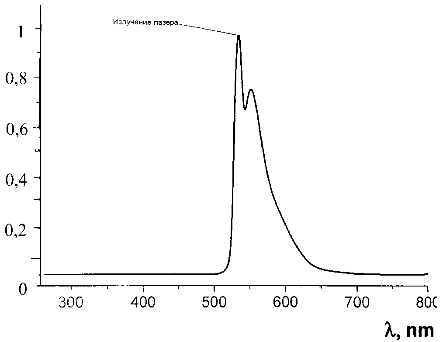
Рисунок 12 - Спектр люминесценции чистого водного раствора эозина G при возбуждении лазером диодной
накачки с λ=532 нм
Максимум люминесценции эозина G наблюдается при 547 нм.
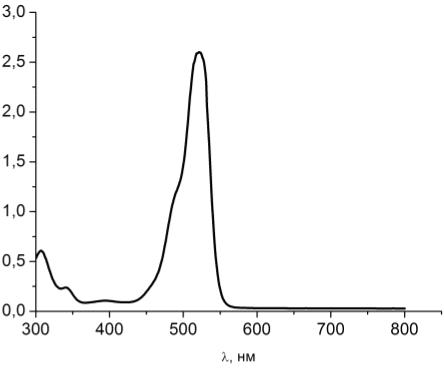
Рисунок 13 - Спектр поглощения чистого водного раствора эозина G
Максимум поглощения эозина G
приходится на 525 нм.
2. Описание экспериментальных установок и методик проведения
экспериментов
2.1
Люминесцентная установка с возможностью откачки воздуха из кюветы с образцом
В рамках данной дипломной работы был проведен эксперимент по установлению
влияния концентраций молекул кислорода на интенсивность люминесценции растворов
красителей с гемоглобином в пористых средах.
Исследования проводились люминесцентно-оптическим методом. Фиксируя
изменение интенсивности люминесценции растворов, можно судить о ее зависимости
от присутствия в молекулах гемоглобина молекулярного кислорода. Схема установки
приведена на рисунке 14.
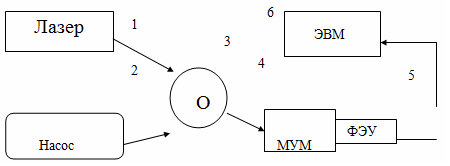
- аппарат лазерный АТС 53-250; 2 - вакуумный насос; 3 - вакуумируемая
кювета с образцом; 4 - монохроматор; 5 - ФЭУ; 6 - ЭВМ.
Рисунок 14 - Схема экспериментальной установки
Возбуждение пористого образца с красителем и гемоглобином осуществлялось
с помощью непрерывного Nd:YAG лазера АТС 53-250, обозначенного 1
на рисунке 14, луч которого попадал на исследуемый образец 3. Свечение образца
попадало на входную щель монохроматора 4. Регистрация сигнала люминесценции
осуществлялась ФЭУ 5, электронные импульсы от которого обрабатывались в компьютере
6. Для питания ФЭУ использовался стабилизатор SH-0105. Напряжение питания составляло 2000 В.
2.1.1 Аппарат
лазерный АТС 53-250
Аппарат предназначен для использования в качестве мощного источника
когерентного излучения в метрологии, голографии, при неразрушающем контроле
поверхностей, шоу программах и т.д. Аппарат может также использоваться в
медицинских учреждения широкого профиля, а также для решения различных
научно-технических задач.
Технические данные аппарата представлены в таблице 2.1.
Таблица 2.1 - Технические данные аппарата лазерного АТС 53-250
|
Наименования параметра
|
Значение
|
|
Длина волны излучения, мкм
|
0,53±0,03
|
|
Диапазон регулирования
выходной мощности лазерного излучения аппарата в непрерывном режиме, Вт
|
от 0 до 0,25
|
|
Модовый состав
|
ТЕМ 00
|
|
Диаметр пучка, мкм
|
<0,5
|
|
Расходимость пучка, мрад
|
<4
|
|
Поляризация
|
линейная 100:1
|
|
Стабильность выходной
мощности (за 1 час) не хуже
|
±5%
|
|
Стабильность положения
светового пятна не хуже, мрад
|
0,5
|
Аппарат состоит из оптического блока и электронного блока управления.
Оптический блок представляет собой оптико-механическую сборку из лазерного
диода с фокусирующими объективами, резонатора твердотельного лазера и систему
поворота лазерного пучка. Блок управления (LDD-10) предназначен для управления режимами работы
оптического блока. Канал управления состоит из трех функциональных частей:
стабилизатор температуры на основе Пельтье-элемента;
блок управления током лазерного диода;
блок защиты лазерного диода.
2.1.2 Вакуумный мини-насос Милливак
Вакуумный мини-насос Милливак предназначен для создания вакуума при
фильтрации воды и водных растворов. Компактный и экономичный источник вакуума
для лабораторных применений.
Насос достигает максимального вакуума 160 мбар (при нулевом потоке
воздуха). Требуется менее 4 минут, для того, чтобы создать вакуум до 200 мбар в
5-литровой емкости.

Рисунок 15 - Внешний вид вакуумного мини-насоса
2.1.3
Монохроматор МДР-204
Монохроматор МДР-204 предназначен для использования в качестве источника
монохроматического излучения в спектральном диапазоне от 190 до 5000 нм.
Встроенный в монохроматор микропроцессорный контроллер обеспечивает
автоматическую установку любой заданной длины волны в рабочем спектральном
диапазоне. Управление сканированием производится от кнопок пульта управления,
значение текущей длины волны высвечивается на встроенном дисплее.
Встроенный микропроцессорный контроллер обеспечивает управление двумя
дополнительными шаговыми двигателями, а также прием и обработку сигналов от
двух внешних приемников излучения. Подключение внешних устройств производится
через разъемы, расположенные на корпусе монохроматора.
Таблица 2.2 - Технические характеристики монохроматора МДР-204
|
Наименование параметра
|
Значение
|
|
Спектральный диапазон, нм
|
190-5000
|
|
Оптическая схема
|
Эберта-Фасти
|
|
Фокусное расстояние
объектива, мм
|
300
|
|
Размеры дифракционной
решетки, мм
|
40х50
|
|
Ширина щелей, мм
|
0-4
|
|
Разрешающая способность
|
12000
|
|
Погрешность установки длины
волны, нм
|
0,5
|
Устройство монохроматора предусматривается возможность непосредственного
подключения к монохроматору внешней ЭВМ через последовательный порт RS232 для управления и регистрации
спектров.
Для работы с внешней ЭВМ используется программное обеспечение, работающее
в среде MSWindows 9X/NT.
2.2 Лазерный кинетический
спектрохронограф с возможностью откачки воздуха из кюветы с образцом
Целью следующего эксперимента, поставленного в данной работе, являлось
установление зависимости кинетики замедленной флуоресценции красителей от
содержания кислорода в гемоглобине пористого образца. В случае наличия
изменений в характере получаемых кинетик при откачке воздуха из кюветы, можно
судить о наличии вышеуказанной зависимости. Схема установки приведена на
рисунке 16.
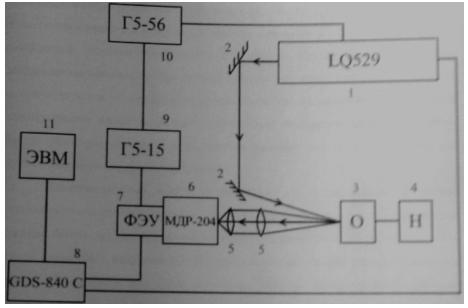
1 - лазер LQ529, 2 -
зеркало; 3 - вакуумируемая кювета с образцом; 4 - вакуумный насос; 5 -
собирающая линза; 6 - монохроматор; 7 - ФЭУ; 8 - осциллограф; 9 - запирающий
генератор Г5-15; 10 - генератор импульсов Г5-56; 11 - компьютер.
Рисунок 16 - Схема экспериментальной установки
Лазерный импульс из лазера 1 отражался от зеркала 2 и попадал на
вакуумируемую кювету с образцом 3. Это приводило к возбуждению молекул
красителя и переходу их в триплетное состояние с последующим испусканием
замедленной флуоресценции. При необходимости, перед проведением эксперимента
давление в кювете понижалось с помощью вакуумного насоса 4. Пройдя через
собирающие линзы 5, сигнал замедленной флуоресценции попадал на щель
монохроматора 6 и усиливался ФЭУ 7. На ФЭУ подавался сигнал запирания лазерного
излучения с генератора 9, а генератор 10 служил для подачи внешних сигналов для
генерации лазерных импульсов. Полученные в результате сигналы выводились на
экран осциллографа 8, который также служил для синхронизации лазерных
импульсов. А также на экран компьютера.
2.2.1 Импульсный лазер LQ529
Лазерный комплекс предназначен для генерации мощных коротких импульсов
когерентного излучения на 1064 нм, 532 нм, 355 нм и 266 нм.
Работа лазера основана на стимулированной эмиссии фотонов электронами,
которые инверсно аккумулируются на верхнем рабочем уровне активной среды лазера
при накачке. Кристаллы Nd3+:YAG используются в качестве активных элементов лазера,
для оптической накачки используются ксеноновые лампы. Активные элементы и лампа
накачки помещены в диффузный отражатель для обеспечения оптимальной передачи
энергии от лампы накачки к активным элементам.
Резонатор лазера содержит ячейку Поккельса с DKDP кристаллом и
поляризационные компоненты. Работа в режиме модулированной добротности
предполагает, что в процессе разряда лампы накачки на элетрооптический затвор
подается "запирающее" напряжение, что приводит к ухудшению
добротности резонатора. При этом генерация излучения отсутствует, энергия
аккумулируется на верхнем энергетическом уровне активной среды. Далее
"запирающее" напряжение ячейки Поккельса снимается, добротность
резонатора резко улучшается и формируется короткий высоко интенсивный импульс
генерации лазерного излучения.
Высокая пиковая мощность импульсов с модулированной добротностью обеспечивает
эффективное преобразование частоты в нелинейных кристаллах. Когда
высокоинтенсивные импульсы излучения (интенсивность которых сравнима с
напряженностью внутриатомных полей) распространяются в нелинейной среде,
нелинейный "отклик" среды - волна поляризации - содержит наряду с
гармониками основной частоты компоненты с суммарными и разностными частотами.
Самое важное условие - наличие фазового синхронизма - предполагает равенство
фазовых скоростей (показателей преломления) для основной и других частот, преобразуемых
в кристалле. Из-за дисперсии в кристаллах, такое равенство возможно только в
двулучепреломляющих кристаллах.
В качестве генератора второй гармоники (длина волны 532 нм) в LQ529 лазере используется кристалл КТР,
имеющий II тип фазового синхронизма.
Для преобразования излучения второй гармоники (длина волны 532 нм) и
непреобразованного в КТР кристалле излучения основной гармоники (длина волны
1064 нм) в излучение третьей/четвертой гармоники (длина волны 355/266 нм) в
лазере LQ529 используются нелинейные
кристаллы.
Таблица 2.3 - Технические характеристики лазера LQ529
|
Наименование параметра
|
Значение
|
|
Длина волны λ, нм
|
1064, 532, 355, 266
|
|
Диаметр пучка на λ=1024 нм, мм
|
6±0,5
|
|
Длительность импульса, нс
|
10-13
|
|
Частота следования
импульсов, Гц
|
10-20
|
|
Энергия импульса, мДж
|
|
λ=1064
нм
|
500
|
|
λ=532
нм
|
280
|
|
λ=355
нм
|
110
|
|
λ=266
нм
|
100-70
|
Рисунок 17 - Внешний вид лазера LQ529
2.2.2
Осциллограф цифровой GDS-840
С
Осциллограф цифровой GDS-840
С предназначен для исследования и измерения параметров периодических сигналов в
полосе частот 0…250 МГц и однократных сигналов в полосе частот до 10 МГц.
Осциллограф способен обеспечить цифровое запоминание, цифровое измерение в
диапазоне амплитуд от 2мВ до 300 В и временных интервалов от 1 нс до 10 с,
автоматическую установку размеров изображения, автоматическое измерение
амплитудно-временных параметров входного сигнала с выводом результата измерения
на экран дисплея.
Тракт вертикального отклонения: пределы допускаемого значения абсолютной
погрешности измерения напряжения в опорном диапазоне частот до 7,5 МГц при
непосредственном входе составляют:
± (0,03 - Uизм + 0,05 • К),
гдеUизм - измеренное значение напряжения, В;
К - величина численно равная установленному значению коэффициента
отклонения.
Таблица 2.4 - Параметры входов каждого из каналов
|
Наименование параметра
|
Значение
|
|
Активное сопротивление при
непосредственном входе, МОм
|
1 ± 2%
|
|
Входная емкость, не более,
Пф
|
22
|
|
C делителем 1:10 активное сопротивление, Мом
|
10 ± 2%
|
|
Сходная емкость, не более,
Пф
|
19
|
Допускаемое суммарное значение постоянного и переменного напряжения на
входе каждого из каналов усилителя не более 300 В, при этом частота переменного
напряжения не должна превышать 1 КГц.
Тракт горизонтального отклонения: пределы допускаемого значения
абсолютной погрешности измерения временных интервалов составляет:
± (0,0001 • Тизм + 0,045 • К + 40пс),
где Тизм - измеренное значение временного интервала, с;
К - величина численно равная установленному значению коэффициента
развертки, с.
Осциллограф обеспечивает автоматический поиск сигнала, автоматическую
установку коэффициента развертки, коэффициента вертикального отклонения и
уровня запуска в полосе частот от 10 Гц до 150 МГц.
Осциллограф обеспечивает возможность записи во внутреннюю память и вызова
15 установок положения органов управления осциллографа при исследовании и
измерении формы входного сигнала.
Осциллограф обеспечивает возможность записи во внутреннюю память и вызова
2 форм сигнала отображаемых на ЭЛТ.
2.3
Спектрофотометр Т70
Спектрофотометр Т70 использовался для получения спектров поглощения растворов
гемоглобина и красителей разных концентраций в воде.
По полученным спектрам поглощения можно было судить о характере
взаимодействия молекул красителя с гемоглобином.
Таблица 2.5 - Технические характеристики спектрофотометра Т70
|
Наименование параметра
|
Значение
|
|
Диапазон длин волн, нм
|
190-1100
|
|
Спектральная полоса
пропускания, нм
|
2
|
|
Точность длины волны, нм
|
±0,3 (с автоматическим
исправлением длины волны)
|
|
Воспроизводимость длин
волн, нм
|
0,2
|
|
Расхождение светового пучка
|
<0,12%Т(220 нм)
|
|
Фотометрический диапазон, Abs
|
-0.3 - 3.0
|
2.4
Электронные лабораторные весы HR-60
Весы лабораторные электронные HR-60 предназначены для статического
измерения массы веществ и материалов и могут применяться в лабораториях
различных предприятий и организаций.
Рисунок 18 - Внешний вид лабораторных электронных весов серии HR
Весы серии HR имеют несколько
функций, позволяющих выполнить настройку на внешние условия:
регулировка скорости отклика, предназначена для получения наиболее
быстрых либо более стабильных показаний;
калибровка с использованием калибровочной гири, позволяющая исключить
возможные погрешности, связанные с силой гравитации, высотой над уровнем моря,
атмосферным давлением и влажностью.
Таблица 2.6 - Технические характеристики лабораторных электронных весов HR-60
|
Наименование параметра
|
Значение
|
|
Наименьший предел
взвешивания, г
|
0,01
|
|
Наибольший предел
взвешивания, г
|
60
|
|
Дискретность отсчета, мг
|
0,1
|
|
Время установления
показаний, не более, с
|
2,5
|
|
Рабочий диапазон температур,
°С
|
5…40
|
|
Рекомендованный
калибровочный вес, г
|
50
|
2.5
Приготовление растворов
Растворы были приготовлены путем разведения сухой порошкообразной
компоненты в растворителе. В качестве растворителя выступала дистиллированная
вода. Необходимая масса компоненты рассчитывались по формуле:
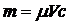 (2.1)
(2.1)
где 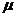 - молекулярный вес вещества;
- молекулярный вес вещества;
с - необходимая концентрация раствора;
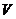 - объем, на который разводится раствор.
- объем, на который разводится раствор.
Для гемоглобина μ=66000 г/моль, для родамина 6G
μ=450,5 г/моль, для
эозина G
μ=692 г/моль, для
эритрозина μ=879,4 г/моль.
Необходимые массы веществ взвешивались на аналитических лабораторных
весах серии HR.
2.6 Методика
заливки пленок
Для проведения кинетического эксперимента с откачкой воздуха из
окрашенных пористых образцов с гемоглобином потребовалось приготовить пленки с
раздробленными пористыми образцами.
В качестве подложки для пленок использовались стекла для микроскопа,
предварительно приведенные к размеру 2,5х2 см. Они очищались от различных
органических и неорганических загрязнений следующим образом.
Стеклянная подложка тщательно промываемся в проточной воде. После этого
подложка промываемся дистиллированной водой. Очищенная таким образом подложка
помещается на несколько минут в ёмкость с этиловым спиртом или ацетоном для
лучшего очищения ее поверхности (обезжиривания). После чего промывается
дистиллированной водой.
По завершении процесса очистки подложка высушивалась потоком теплого воздуха.
Далее на подложку наносился слой (около 1 мм) раздробленных пористых
образцов (силикагеля), предварительно окрашенных в растворе красителя и
гемоглобина. Для закрепления слоя раздробленных образцов наносился 1% раствор
ПВБ.
2.7
Проведение экспериментов
На спектрофотометре Т70 были исследованы жидкие растворы гемоглобина
(концентрация С=10-7 моль/л), органических красителей родамина 6G (концентрацией С=10-6
моль/л), эозина G (концентрацией
С=10-6 моль/л) и эритрозина (концентрацией С=10-6 моль/л)
и смеси этих красителей с гемоглобином в указанных концентрациях. По сдвигу
спектров поглощения можно судить о характере взаимодействия молекул того или
иного красителя с молекулами гемоглобина.
Далее было решено провести серию экспериментов на люминесцентной
установке, изображенной на рисунке 2.1. Вакуумный насос в этой серии
экспериментов не использовался. Были использованы следующие растворы красителей
с гемоглобином: родамин 6G с
гемоглобином (10-4 моль/л и 10-5 моль/л соответственно),
эритрозин с гемоглобином (10-4 моль/л и 10-5 моль/л
соответственно) и эозин G с
гемоглобином (10-4 моль/л и 10-5 моль/л соответственно).
Была исследована зависимость люминесценции красителей от относительного
содержания красителя и гемоглобина в растворе. Исследуемый раствор помещался в
кварцевую кювету, закрепляемую на вращающемся основании. Воздействие лазерного
излучения на кювету с раствором приводило к возбуждению молекул красителя.
Сигнал люминесценции красителя, пройдя через монохроматор, попадал в ФЭУ и,
пройдя через ряд преобразователей, выводился на мониторе компьютера.
Компьютером регистрировалась усредненная кривая. Проводилось снятие серии
сигналов с различными красителями, указанными выше, для усреднения результата.
Для каждого раствора снималось от трех до семи кривых сигнала люминесценции.
Далее данные усреднялись, и полученные значения параметров обрабатывались в
пакете Origin. По прошествии 24 часов данный
эксперимент повторялся с этими же растворами красителей и гемоглобина для
выявления воздействия кислорода и оседании гемоглобина на молекулах красителя.
В дальнейшем были проведены эксперименты по исследованию зависимости
люминесценции красителей в пористых средах (силикагели), окрашенных раствором
красителей и гемоглобина. Проведены 2 серии экспериментов: с полностью
высушенными пористыми средами и чуть влажными, также в эксперименте участвовало
2 вида лазеров: синий и зеленый, для выявления степени точности полученных
результатов. Эксперимент проводился на люминесцентной установке с возможностью
откачки воздуха из кюветы с образцом, изображенной на рисунке 14. Концентрация
красителя составляла 10-3, 10-4 моль/л, а концентрация
гемоглобина 10-5 моль/л.
Для снижения концентрации кислорода в гемоглобине, образцы помещались в
вакуумируемую кювету. С помощью вакуумного насоса давление в кювете понижалось
до 0,8 мбар. Дело в том, что, согласно теории, при снижении давления над
раствором гемоглобина, происходит его (гемоглобина) дезоксигенация, то есть
молекулы кислорода, связанные гемоглобином, покидают его. Снимались серии
кривых без откачки воздуха из кюветы и с откачкой.
Результаты экспериментов представлены в главе 3.
3. Результаты
экспериментов
3.1
Абсорбционный эксперимент
Как уже отмечалось в главе 2, в рамках данной дипломной работы был
проведен эксперимент по исследованию взаимодействия молекул красителя с
молекулами гемоглобина. Эксперимент проводился методом абсорбционной
спектроскопии. Концентрация гемоглобина в исследуемых растворах составляла 10-7
моль/л, концентрации красителей в растворах составляли 10-6 моль/л.
Исследования проводились на спектрофотометре Т70.
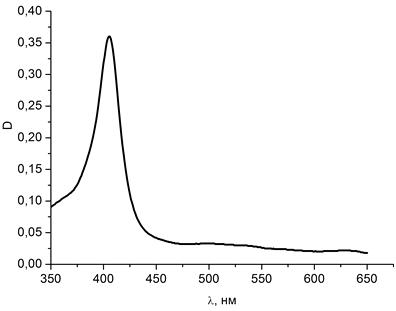
Рисунок 19 - Спектр поглощения водного раствора гемоглобина (концентрации
10-7 моль/л) при н.у.
На рисунке 19 представлен спектр поглощения водного раствора гемоглобина
концентрации С=10-7 моль/л. На длине волны, равной 405 нм, имеется
характерный для гемоглобина пик поглощения.
Было выдвинуто предположение, что в результате высаживание молекул
красителя на молекулу гемоглобина в растворе, содержащем гемоглобин и
краситель, произойдет изменение спектра поглощения красителя, а именно сдвиг
максимума поглощения.
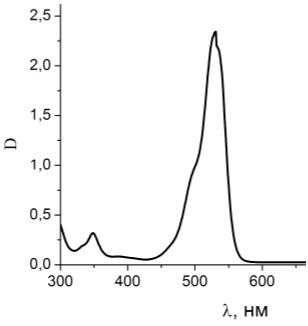
Рисунок 20 - Спектр поглощения чистого раствора родамина 6G
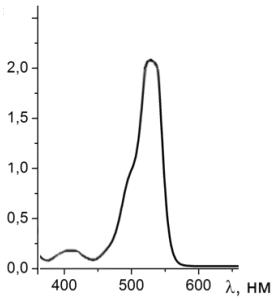
Рисунок 21 - Спектр поглощения раствора родамина 6G с гемоглобином
Как видно из спектров поглощения на рисунке 20 и 21, пик поглощения
родамина 6G при добавлении в раствор гемоглобина
не смещается, уменьшается интенсивность поглощения за счет присутствия
гемоглобина. На графике виден пик гемоглобина на λ=405 нм.
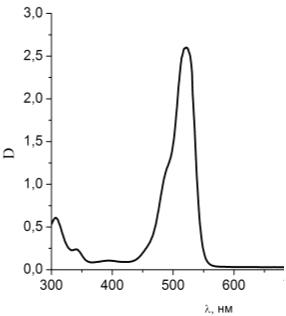
Рисунок 22 - Спектр поглощения чистого раствора эозина G
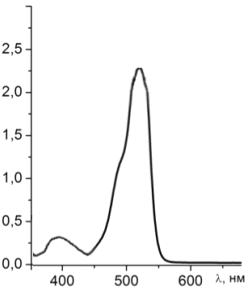
Рисунок 23 - Спектр поглощения раствора эозина G с гемоглобином
Проводя анализ рисунков 22 и 23 сравним спектр поглощения чистого
раствора эозина G и раствора
эозина G при добавлении в него гемоглобина.
Пик поглощения эозина G при
добавлении в раствор гемоглобина не смещается, уменьшается интенсивность
поглощения за счет присутствия гемоглобина. На графике также виден пик
гемоглобина на λ=405 нм.
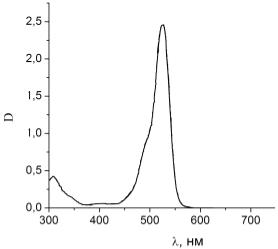
Рисунок 24 - Спектр поглощения чистого раствора эритрозина
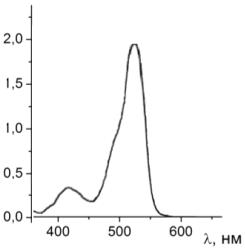
Рисунок 25 - Спектр поглощения раствора эритрозина с гемоглобином
Из анализа рисунков 24 и 25 также видно, что пик поглощения раствора
эритрозина, как и в случае с родамином 6G, уменьшается при добавлении в раствор гемоглобина, смещение
не происходит, а интенсивность поглощения уменьшается за счет присутствия
гемоглобина.
Абсорбционный эксперимент определяет характер взаимодействия красителей с
гемоглобином. У всех красителей при добавлении в раствор гемоглобина снижается
интенсивность поглощения, за счет присутствия гемоглобина.
3.2
Люминесцентный эксперимент с жидкими растворами красителей и гемоглобина
Так как результаты абсорбционного эксперимента не дали ответа на вопрос о
взаимодействии красителей с гемоглобином, было решено провести серию
люминесцентных экспериментов с жидкими растворами красителей с добавлением
гемоглобина с разницей в проведении экспериментов в 24 часа.
Исследования проводились на установке, описанной в 2.1, без участия
вакуумного насоса. Проводился эксперимент с растворами красителей с
концентрациями: родамин 6G с
гемоглобином (10-4 моль/л и 10-5 моль/л соответственно),
эритрозин с гемоглобином (10-4 моль/л и 10-5 моль/л
соответственно) и эозин G с
гемоглобином (10-4 моль/л и 10-5 моль/л соответственно).
Сдвиг спектра люминесценции раствора красителя и гемоглобина, полученный
на 24 часа позже первого эксперимента, свидетельствовал бы о взаимодействии
молекул красителя с молекулами гемоглобина.
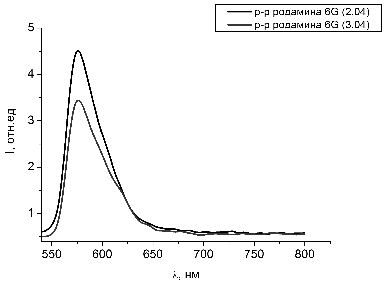
Рисунок 26 - Спектры люминесценции растворов родамина 6G с добавлением гемоглобина с разницей
в 24 часа

Рисунок 27 - Спектр люминесценции чистого раствора родамина 6G
На рисунке 26 показано временное изменение спектров люминесценции водных
растворов гемоглобина и красителя родамина 6G. В сравнение на рисунке 27 приводится спектр люминесценции
чистого раствора родамина 6G.
Сдвига пика спектра люминесценции не наблюдается. Мы предполагаем, что снижение
интенсивности люминесценции растворов происходит за счет выгорания красителя
под действием лазерного излучения.
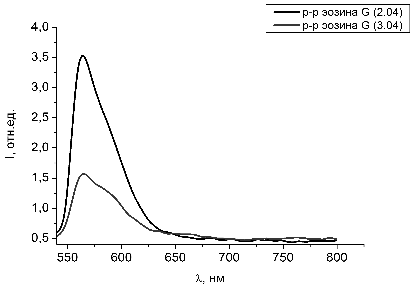
Рисунок 28 - Спектры люминесценции растворов эозина G с добавлением гемоглобина с разницей
в 24 часа
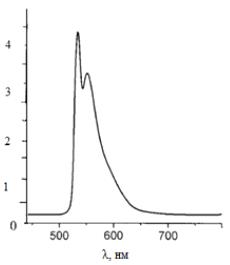
Рисунок 29 - Спектр люминесценции чистого раствора эозина 6G
Как видно из рисунков 28 и 29 сдвига пиков люминесценции у красителя
эозина G при добавлении гемоглобина и с
течением времени не наблюдается. Происходит только снижение интенсивности
люминесценции за счет выгорания красителя в растворах под действием лазерного
излучения.
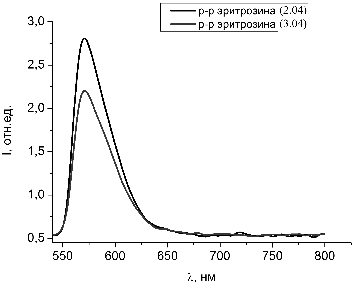
Рисунок 30 - Спектры люминесценции растворов эритрозина с добавлением
гемоглобина с разницей в 24 часа
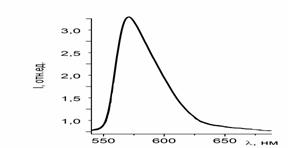
Рисунок 31 - Спектр люминесценции чистого раствора эозина 6G
Представленные на рисунке 30 и 31 спектры люминесценции свидетельствуют
об уменьшении интенсивности люминесценции растворов с течением времени и с
добавление к раствору гемоглобина. Также сделано предположение, что происходит
это за счет выгорания красителя под действием лазерного излучения. Процессы
взаимодействия гемоглобина с красителем достаточно длительные, требуется время
для образования некого комплекса.
.3 Статистический анализ люминесценции пористых образцов, окрашенных
раствором красителей и гемоглобина
гемоглобин кислород люминесценция абсорбционный
Для дальнейшего исследования взаимодействия красителей с гемоглобином
было решено провести серию люминесцентных экспериментов с пористыми образцами
(силикагелем), окрашенными раствором красителей и гемоглобина в различных
концентрациях (10-4, 10-3 моль/л и 10-5 моль/л
соответственно). Окрашивание проводилось в течение 1-1,5 минут путем помещение
выбранного сухого образца в заранее приготовленный раствор. Эксперимент
проводился на установке, описанной в 2.1, без откачки кислорода. Целью данного
эксперимента являлось выявление влияния молекул гемоглобина на изменение пика
люминесценции окрашенных пористых образцов.
Для каждого из растворов красителя с гемоглобином были подготовлены
несколько образцов для сбора статистических данных по эксперименту с целью
нахождения среднего значения люминесценции, дисперсии и истинного значения.
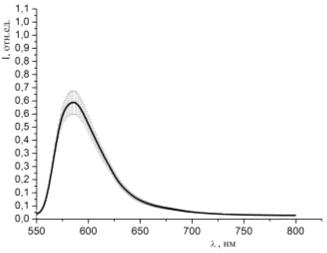
Рисунок 32 - Спектр люминесценции образцов, окрашенных раствором родамина
6G (С=10-3 моль/л) и
гемоглобина (10-5 моль/л) при освещении зеленым лазером (534 нм)
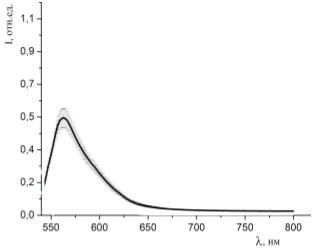
Рисунок 33 - Спектр люминесценции образцов, окрашенных раствором родамина
6G (С=10-4 моль/л) и
гемоглобина (10-5 моль/л) при освещении зеленым лазером (532 нм)

Рисунок 33 - Спектр люминесценции образцов, окрашенных раствором эозина G (С=10-3 моль/л) и
гемоглобина (10-5 моль/л) при освещении зеленым лазером (532 нм)
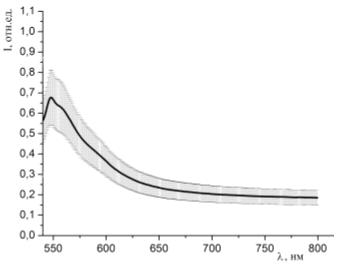
Рисунок 35 - Спектр люминесценции образцов, окрашенных раствором эозина G (С=10-4 моль/л) и
гемоглобина (10-5 моль/л) при освещении зеленым лазером (532 нм)
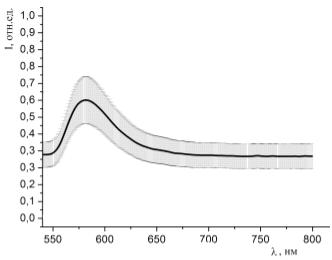
Рисунок 36 - Спектр люминесценции образцов, окрашенных раствором
эритрозина (С=10-3 моль/л) и гемоглобина (10-5 моль/л)
при освещении зеленым лазером (532 нм)
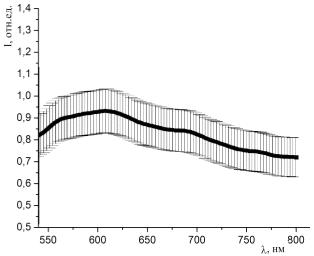
Рисунок 37 - Спектр люминесценции образцов, окрашенных раствором
эритрозина (С=10-4 моль/л) и гемоглобина (10-5 моль/л)
при освещении зеленым лазером (532 нм)
Также был проведен ряд таких же экспериментов, но с воздействием синего
лазера
(λ=445 нм).
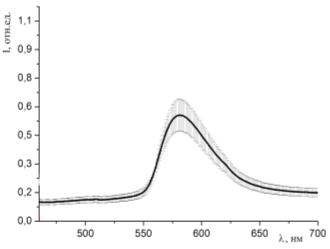
Рисунок 38 - Спектр люминесценции образцов, окрашенных раствором родамина
6G (С=10-3 моль/л) и
гемоглобина (10-5 моль/л) при освещении синим лазером (445 нм)
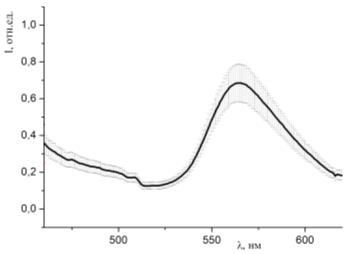
Рисунок 39 - Спектр люминесценции образцов, окрашенных раствором родамина
6G (С=10-4 моль/л) и
гемоглобина (10-5 моль/л) при освещении синим лазером (445 нм)
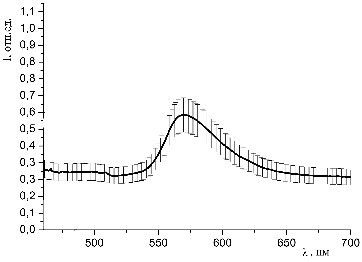
Рисунок 40 - Спектр люминесценции образцов, окрашенных раствором эозина G (С=10-3 моль/л) и
гемоглобина (10-5 моль/л) при освещении синим лазером (445 нм)
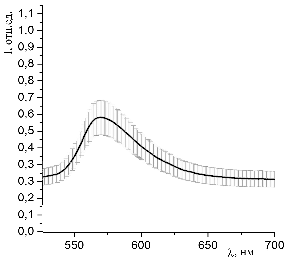
Рисунок 41 - Спектр люминесценции образцов, окрашенных раствором эозина G (С=10-4 моль/л) и
гемоглобина (10-5 моль/л) при освещении синим лазером (445 нм)
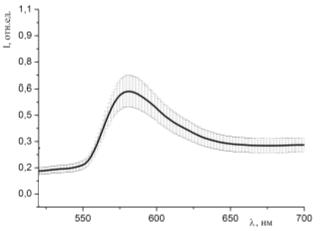
Рисунок 42 - Спектр люминесценции образцов, окрашенных раствором
эритрозина (С=10-3 моль/л) и гемоглобина (10-5 моль/л)
при освещении синим лазером (445 нм)
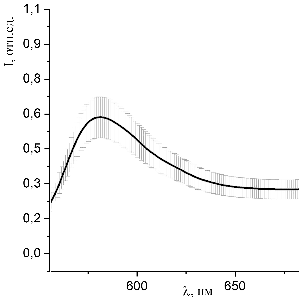
Рисунок 43 - Спектр люминесценции образцов, окрашенных раствором
эритрозина (С=10-4 моль/л) и гемоглобина (10-5 моль/л)
при освещении синим лазером (445 нм)
Представленные на рисунках 32 - 43 спектры люминесценции образцов,
окрашенных различными красителями с различными концентрациями с добавлением в
раствор гемоглобина, говорят о том, что интенсивность люминесценции понижается
при уменьшении концентрации красителя в исследуемом растворе.
По полученным данным однозначно можно судить о наличии взаимодействия
молекул красителей с молекулами гемоглобина в пористых средах.
3.4
Люминесцентный эксперимент с пористыми образцами, окрашенными раствором
красителей и гемоглобина с откачкой воздуха
Далее была проведена серия таких же экспериментов, с участием вакуумного
насоса, то есть с откачкой воздуха из образцов. Целью данного эксперимента
являлось выяснение степени влияния молекулярного кислорода, содержащегося в
гемоглобине, на интенсивность люминесценции красителей.
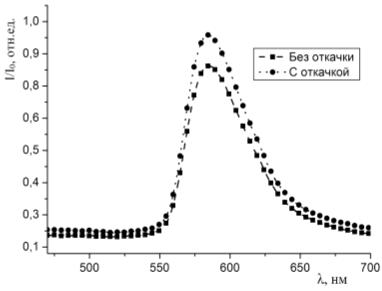
Рисунок 44 - Зависимость спектра люминесценции пористого образца,
окрашенного раствором родамина 6G
(С=10-3 моль/л) с гемоглобином от давления в кювете с образцом
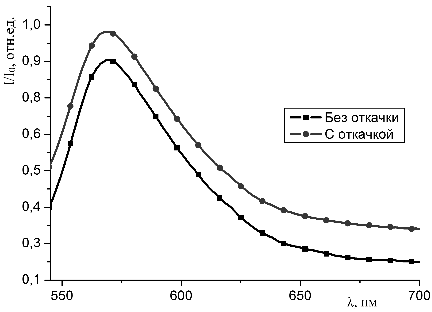
Рисунок 45 - Зависимость спектра люминесценции пористого образца,
окрашенного раствором родамина 6G
(С=10-4 моль/л) с гемоглобином от давления в кювете с образцом
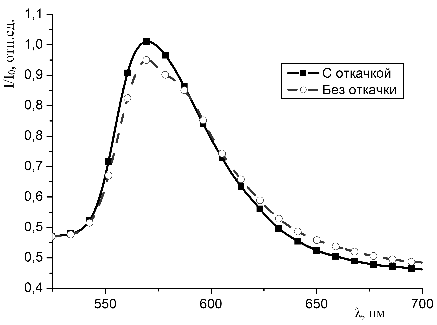
Рисунок 46 - Зависимость спектра люминесценции пористого образца,
окрашенного раствором эозина G
(С=10-3 моль/л) с гемоглобином от давления в кювете с образцом
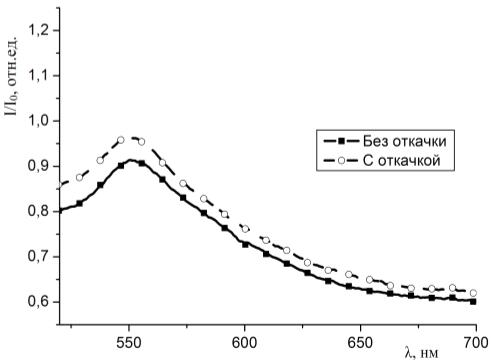
Рисунок 47 - Зависимость спектра люминесценции пористого образца,
окрашенного раствором эозина G
(С=10-4 моль/л) с гемоглобином от давления в кювете с образцом
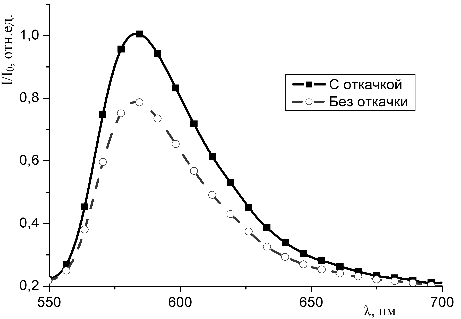
Рисунок 48 - Зависимость спектра люминесценции пористого образца, окрашенного
раствором эритрозина (С=10-3 моль/л) с гемоглобином от давления в
кювете с образцом
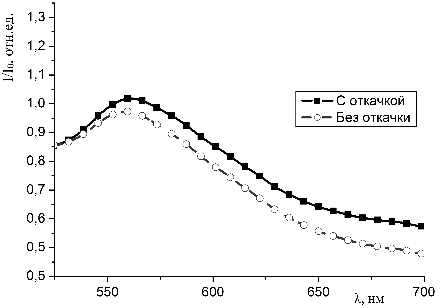
Рисунок 49 - Зависимость спектра люминесценции пористого образца,
окрашенного раствором эритрозина (С=10-4 моль/л) с гемоглобином от
давления в кювете с образцом
Из рисунков 44 - 49 видно, что при откачке воздуха из кюветы с образцом и
снижении давления до 1 мбар происходит увеличение интенсивности люминесценции
красителя. Концентрация родамина 6G 10-3 и 10-4 моль/л, концентрация гемоглобина - 10-5
моль/л.
Так как в данном эксперименте у всех исследуемых красителей в разных
концентрациях обнаружена идентичная зависимость спектра люминесценции от
давления в кювете с образцом, можно сделать утверждение о том, что молекулы
красителей взаимодействуют с молекулами гемоглобина.
3.5
Кинетический эксперимент с пленками силикагеля, окрашенного растворами
красителей с гемоглобином и покрытого ПВБ
После получения данных о зависимости люминесценции пористых образцов,
окрашенных раствором красителей и гемоглобина, от давления в кювете с образцом
было принято решение исследовать зависимость замедленной флуоресценции твердых
растворов красителей с гемоглобином от давления в кювете с образцом.
Эксперимент проводился на установке, описанной в 2.2, пленки с раздробленным
силикагелем заливались по технологии, описанной в 2.6. В качестве красителей
использовались эозин G,
эритрозин и родамин 6G в
концентрации 10-3 моль/л каждый. Концентрация гемоглобина составляла
10-5 моль/л.
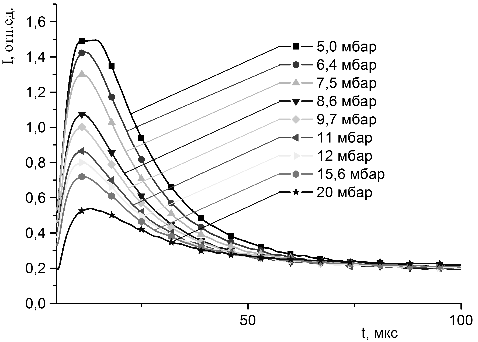
Рисунок 50 - Зависимость сигнала замедленной флуоресценции эритрозина от
давления в кювете с образцом
Таблица 3.1 - Точки максимума при различных значениях давления для
красителя эритрозина
|
P, мбар
|
Тmax
|
|
5,0
|
1,49
|
|
6,4
|
1,43
|
|
7,5
|
1,3
|
|
8,6
|
1,07
|
|
9,7
|
1,0
|
|
11
|
0,86
|
|
12
|
0,8
|
|
15,6
|
0,72
|
|
20
|
0,54
|
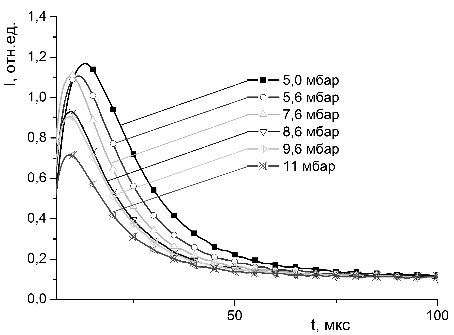
Рисунок 51 - Зависимость сигнала замедленной флуоресценции эозина G от давления в кювете с образцом
Таблица 3.2 - Точки максимума при различных значениях давления для
красителя эозина G
|
P, мбарТmax
|
|
|
5,0
|
1,17
|
|
5,6
|
1,1
|
|
7,6
|
1,11
|
|
8,6
|
0,93
|
|
9,6
|
0,9
|
|
11
|
0,71
|

Рисунок 52 - Зависимость сигнала замедленной флуоресценции родамина 6G от давления в кювете с образцом
Таблица 3.3 - Точки максимума при различных значениях давления для
красителя родамина 6G
|
P, мбарТmax
|
|
|
5,4
|
0,81
|
|
5,9
|
0,76
|
|
6,9
|
0,73
|
|
7,9
|
0,73
|
|
9
|
0,63
|
|
15
|
0,57
|
Представленные на рисунках 50 - 52 кинетики замедленной флуоресценции
различных красителей наглядно демонстрируют, что при минимальных концентрациях
кислорода наблюдаются максимумы интенсивности сигнала. Кривые имеют ярко
выраженный горб, с последующим спадом интенсивности. Кроме интенсивности по
графикам можно наблюдать, что сдвигается время накопления на максимум.
Основные результаты и выводы
В рамках данной дипломной работы было проведено исследование
взаимодействия молекул красителя с молекулами гемоглобина. В частности:
В результате взаимодействия красителей с гемоглобином происходит снижение
интенсивности люминесценции и снижение пиков поглощения растворов на 25%.
Влияние пористых структур и концентраций кислорода на люминесценцию
растворов красителей и гемоглобина приводит к усилению люминесценции до 30%.
Наблюдается высокая зависимость интенсивности замедленной флуоресценции
от концентрации кислорода в кювете с пленками пористых сред, окрашенных
раствором красителя и гемоглобина.
Результаты исследований, проведенных в рамках данной работы, позволяют
сделать предположение о том, что молекулы красителей взаимодействуют с
молекулами гемоглобина в растворах и пористых средах, окрашенных раствором
красителей и гемоглобина, а также параметры пористых сред оказывают влияние на
характер взаимодействия.
Заключение
В результате проведенных экспериментов была выделена зависимость
люминесценции от концентрации кислорода в растворах красителей с гемоглобином
или пористых образцах, окрашенных растворами красителей с гемоглобином.
Полученные исследования помогут тщательнее изучить влияние молекулярного
кислорода на гемоглобин крови. Результаты данных исследований можно использовать
для диагностики и лечения, для изучения отдельных органов человека и животных.
Необходимо продолжать исследования в данной области, в силу важности
кислорода в процессах жизнедеятельности всего организма.
В заключение хочу выразить свою благодарность Центру лазерной и
информационной биофизики ОГУ за консультацию и помощь в проведении
экспериментальных исследований моей выпускной квалификационной работы.
Список использованных источников
Биологические структуры и функции на молекулярном уровне:
сб.науч.тр. / АН СССР. - М.: изд-во АнСССР, 1962. - 124 с.
Лауреаты Нобелевской премии: энциклопедия. - М.: Прогресс,
1992. - 256 с.
Перуц, М. Молекулярный механизм дыхания / М. Перуц // Наука и
человечество. - М.: Знание, 1966. - С. 50-59, С. - 85-86.
Скулачев, В.П. Кислород в живой клетке: добро или зло / В.П.
Скулачев. - М. Биология, 199. - 312 с.
Онлайн - справочник химических элементов [Электронный
ресурс]. - Режим доступа: http://webelements.narod.ru/elements/O.htm.
Сайт о дыхании. [Электронный ресурс] -
Режим доступа: http://www.chakrachka.ru/dyhanie/theory/kislorod.htm.
Чарный, А.М. Патофизиология гипоксических состояний / А.М.
Чарный. - М.: Медгиз, 1961. - 342 с.
Кучеренко, М.Г. Изменение кинетики аннигиляционной
люминесценции красителей в полимерах под действием лазерного импульса / М.Г.
Кучеренко, М.П. Мельник, Г.М. Кецле, С.Н. Летута // Оптика и спектроскопия. -
1995. - №4. - С. 649-653.
Bolton, P.H., Kenner,
R.D., Khan, A.U. // J. Chem. Phys. 1972. - V. 57. - №12. - Р. 5604-5605.
Кецле, Г.А., Кучеренко, М.Г., Якупов, Р.М. // Оптика и
спектроскопия. - 1989. - Т. 67. - В. 5. - С. 1090-1094.
. Кучеренко, М.Г., Мельник, М.П., Якупов, Р.М. // Известия АН
СССР. Сер. Физ. 1990. - Т. 54. - №3. - С. 489-495.
Кецле, Г.М., Кучеренко, М.Г., Якупов, Р.М. // Оптика и спектроскопия.
- 1989. - Т. 67. - В. 5. - С. 1090-1094.
Кецле, Г.А., Левшин, Л.В., Летута, С.Н. // Оптика и
спектроскопия. - 1990. - Т. - 68. - В. 2. - С. 334.
Кучеренко, М.Г. Селективное лазерное усиление флуктуаций
скорости реакций / Кучеренко, М.Г., Мельник М.П., Кецле, Г.А., Летута, С.Н. //
Известия АН России. Сер. Физ. - 1993. - Т. 57. - №12. - С. 175-180.
Википедия - свободная энциклопедия. [Электронный ресурс] -
Режим доступа: http://ru.wikipedia.org/wiki/Гемоглобин.
Шайтан, К.В. Диффузия лигандов в белках / К.В. Шайтан //
Соросовский образовательный журнал. - 2000. - №6. - С. 8-13.
Волькенштейн, М.В. Биофизика: учеб. руководство / М.В.
Волькенштейн. - М.: Наука, 1988. - 592 с.
Шайтан, К.В. Диффузия лигандов в белках / К.В. Шайтан //
Соросовский образовательный журнал. - 2000. - №6. - С. 13-20.
Шайтан, К.В. К теории миграции лигандов в биомакромолекулах /
К.В. Шайтан, И.В. Упоров, А.Б. Рубин // Молекулярная Биология. - 1985. - Т. 19.
- С. 742-750.
Ягтман, Д.Р. Силикагели / Д.Р. Ягтман. - М.: Химия, 2012. -
472 с.
Кельцев, Н.В. Основы адсорбционной техники / Н.В. Кельцев. -
М., 1984. - 534 с.
Неймарк, И.Е. Силикагель, его получение, свойства и
применение / И.Е. Неймарк, Р.Ю. Шейнфайн. - М.: Химия, 1972. - 200 с.
Механика жидкости и газа // Вестник Нижегородского
университета им. Н.И.Лобачевскогою. - 2011 - №4 (3). - С.777-778.
Головина, А.П. Химический люминесцентный анализ
неорганических веществ / А.П. Головина, Л.В. Левшин. - М.: Химия, 1978. - 248
с.
Левшин, В.Л. Фотолюминесценция жидких и твердых веществ. -
М., 1951. - 378 с.
Асимов, М.А. Стимулирование аэробного метаболизма клеток
низкоинтенсивным лазерным излучением / М.М. Асимов, Р.М. Асимов, А.Н. Рубинов,
С.А. Мамилов, Ю.С. Плаксий // Лазерная медицина. - 2007. - Т. 11. - В. 2. - С.
53.
27 Sychra, V. Atomic fluorescence spectroscopy / V.
Sychra, V. Svoboda, I. Rubeska. - London: Van Nostrand Reinhold co., 1975. -
379 р.
28 Левшин, В.Л. Труды ФИАН. - 1963. - Т 23. - С. 64
Лосев, Н.Ф. Количественный рентгеноспектральный
флуоресцентный анализ / Н.Ф. Лосев. - М.: Наука, 1969. - 366 с.
Верещагин, И.К. Электролюминесценция кристаллов /
И.К.Верещагин. - М.: Наука, 1974. - 279 с.
Вавилов, С.И. Сочинения: в 5 т. / С.И. Вавилов. - М.:
Издательство АН СССР, 1954. - Т. 1. - 344 с.
Антонов - Романовекий, В.В. Кинетика фотолюминесценции
кристаллофосфоров / В.В. Антонов-Романовекий. - М.: Наука, 1966. - 125 с.
33 Montero, E. Spectral pH dependence of erythrosine
in sol-gel silica coating and buffered solutions / E. Montero, M.A. Garcia,
M.A. Villegas. J.Llopis. - 2008. - V. 47. - Р. 1-6.
Pravinata, L.C. Erythrosin B Phosphorescence Monitors
Molecular Mobility and Dynamic Site Heterogeneity in Amorphous Sucrose / Linda
C. Pravinata; Yumin You // Biophysical Journal. - 2005. - V. 88. - Р. 3551-3561.