Современное состояние зообентоса реки Бейсуг и его значение в питании массовых видов рыб
МИНИСТЕРСТВО ОБРАЗОВАНИЯ И НАУКИ
РОССИЙСКОЙ ФЕДЕРАЦИИ
Федеральное государственное бюджетное
образовательное учреждение высшего профессионального образования
"КУБАНСКИЙ ГОСУДАРСТВЕННЫЙ
УНИВЕРСИТЕТ"
(ФГБОУ ВПО "КубГУ")
Кафедра водных биоресурсов и
аквакультуры
ДОПУСТИТЬ К ЗАЩИТЕ В ГЭК
ВЫПУСКНАЯ КВАЛИФИКАЦИОННАЯ РАБОТА
БАКАЛАВРА
Современное состояние зообентоса реки
Бейсуг и его значение в питании массовых видов рыб
Работу выполнила Я.Ю. Нещадим
Факультет биологический
Направление 35.03.08 Водные биоресурсы и аквакультура
Научный руководитель
канд. биол. наук,
заведующий кафедрой А.Н. Пашков
Краснодар 2015
Реферат
Выпускная квалификационная работа бакалавра: ___с.
ЗООБЕНТОС, РЕКА БЕЙСУГ, ЧИСЛЕННОСТЬ, БИОМАССА, ЭКОЛОГИЧЕСКОЕ
СОСТОЯНИЕ, ВИДОВОЙ СОСТАВ.
Объектом исследования послужили пробы зообентоса степной реки
Бейсуг, отобранные в летний и осенний периоды 2014 г.
Целью данной выпускной квалификационной работы являлось
изучение и описание современного состояния зообентосного сообщества реки Бейсуг
и выявление роли зообентосных организмов в питании массовых видов рыб этой
реки.
В данной выпускной квалификационной работе описан
таксономический состав зообентосного сообщества реки Бейсуг и приведены данные
о его численности и биомассе. Изучены особенности пространственного
распределения зообентоса вдоль продольного профиля реки. Рассмотрена динамика
зообентоса по месяцам и в зависимости от района исследования. Проведена оценка
современного состояния зообентоса р. Бейсуг и определена его роль в питании массовых
видов рыб реки.
Содержание
Введение
1. Аналитический обзор
1.1 Общая характеристика зообентоса как экологической группы
гидробионтов
1.2 Таксономический состав и морфо-биологические особенности
речного зообентоса
1.3 Особенности развития зообентоса степных рек
1.4 Антропогенное влияние на зообентос степных рек
2. Физико-географическая характеристика реки Бейсуг
3. Материал и методы исследования
4. Современное состояние зообентоса реки Бейсуг и его значение в
питании массовых видов рыб
4.1 Видовой состав зообентоса
4.2 Пространственная динамика количественных характеристик
зообентоса реки Бейсуг
4.3 Пространственная динамика биомассы зообентоса реки Бейсуг
4.4 Сезонная динамика развития зообентоса реки Бейсуг
4.5 Оценка экологического состояния зообентоса реки Бейсуг
4.6 Значение зообентоса в питании массовых видов рыб
Заключение
Библиографический список
Введение
Территория Краснодарского края обводнена тремя системами рек:
рекой Кубанью и реками её бассейна, черноморскими реками и реками Азово-Кубанской
низменности, называемыми также степными [Борисов, 2005].
К числу степных относят реки, текущие севернее р. Кубани и
впадающие либо непосредственно в Азовское море, либо заканчивающиеся сетью
заболоченных лиманов. Наиболее крупные из них: Ея, Челбас, Бейсуг, Кирпили,
Понура, Сенгели, Ясени, Албаши, Средний Зеленчук [Борисов, 2005].
Человек особенно, в последние 20-30 лет, практически перестал
задумываться над судьбой степных рек, резко изменяя их режим через
строительство дамб, сбрасывая в них животноводческие и бытовые стоки и т.д. В
средней части многие реки превращены в каскад плотин, водохранилищ и прудов.
Как решать вопросы регулирования стока рек, снижения их загрязнения и
использования искусственных водоемов, снижения их негативного влияния на природу
и жизнь человека - важнейшая современная проблема [Белюченко, 2005].
Степные реки в крае играют особую роль. C одной стороны, они
имеют местное значение, с другой - любая отдельная река - это часть общего
водного бассейна. Правильное использование водных ресурсов степной зоны -
будущее этого региона. Погибнут степные реки, и мы лишимся огромной территории,
производящей зерно, потеряем плодороднейшие почвы [Белюченко, 2004].
Густота речной сети степных рек невелика (до 0,3 км/км2).
Они маловодны. Половодье на них наблюдается весной, а в июле - сентябре большинство рек
сильно мелеет, местами пересыхает, образуя мелководные разобщённые плёсы,
интенсивно зарастающие растительностью [Борисов, 1978; Нагалевский, Щеглова,
1993].
Жизнь любой реки неотделима от той территории, по которой она
протекает, и все проходящие в бассейне изменения, отражаются на её состоянии.
Всего 50 лет назад водность любой степной реки в крае была значительно большей,
чем сейчас. Даже такие реки, как Челбас и Кирпили, доносили свои воды до
Азовского моря [Белюченко, 2004].
Современное состояние экосистем степных рек и тенденции его
изменения изучены недостаточно. После комплексной экспедиции биологического
факультета, посвящённой исследованию данной проблемы, результаты которой были
изложены в соответствующем сборнике [Актуальные вопросы., 1992], степные реки
практически не изучались.
За последние 20 лет было опубликовано лишь несколько работ из
данной области. Ряд публикаций Г.В. Волошиной [2006 а, 2006 б, 2007] посвящены
оценке экологического состояния р. Понура. Несколько работ сотрудников
Краснодарского филиала ФГУП "ВНИРО" посвящено описанию показателей
развития кормовой базы рыб в некоторых реках региона [Рыбохозяйственное
освоение…, 2013; Болкунов, Рудакова, Ерзиков, 2014].
При оценке состояния экосистем рек особое значение имеет
использование донных зообентосных сообществ, что неоднократно подчёркивалось
многими исследователями, в отечественной литературе [Москул, 1985, 1987; Абаев,
1992 и др.]. Её анализ показывает, что основой понимания состояния и
функционирования пресноводных экосистем во многом являются данные о
распределении, динамике и структуре донных сообществ.
Оценка состояния рек, в том числе и степных, по показателям
зообентоса имеет преимущество перед иными подходами, которое заключается в том,
что видовой состав и структура донных сообществ отражают состояние экосистемы
за относительно длительный период, тогда как, например, планктонные сообщества
характеризуют качество водоёма в данный момент времени, адекватно реагируя на
кратковременные антропогенные воздействия [Белюченко, 2005].
Необходимость изучения экосистем степных рек обусловлена
высокой степенью их зарегулированности, в силу чего их гидробиологический режим
сильно изменяется, что сказывается на всех звеньях пищевых цепей [Батурин,
1994]. Макрозообентос степных рек изучен очень слабо, а поскольку его
составляющие играют большую роль в процессах очищения водоёма и представляют
собой часть кормовой базы многих рыб, то их изучение, безусловно, важно
[Белюченко, 2005].
Анализ функционирования экосистем малых рек, в силу некоторых
методических сложностей, связанных с динамичностью водотоков, невозможен без
изучения фауны. Фаунистические исследования являются необходимым звеном в
комплексном исследовании оценки экосистем малых рек в условиях изменившегося
режима и для получения сведений о качестве воды рек [Алимов, 1976].
Целью данной выпускной квалификационной работы являлось
изучение и описание современного состояния зообентосного сообщества реки Бейсуг
и выявление роли зообентосных организмов в питании массовых видов рыб этой
реки.
Для достижения цели решали следующие задачи:
. Изучить таксономический состав зообентосного сообщества
реки Бейсуг;
. Получить данные о численности и биомассе зообентоса реки
Бейсуг;
. Описать особенности пространственного распределения
зообентоса изученного водотока вдоль его продольного профиля;
. Выявить сезонную динамику развития зообентоса реки Бейсуг;
. Оценить современное состояние зообентоса р. Бейсуг;
. Определить значение зообентоса в питании массовых видов рыб
реки Бейсуг.
зообентос река бейсуг рыба
1.
Аналитический обзор
1.1 Общая
характеристика зообентоса как экологической группы гидробионтов
Обитатели водной среды - гидробионты, населяют практически
всю гидросферу от её поверхности до максимальных глубин. Значительная часть
гидробионтов обитает на дне или на другом субстрате - растениях, подводных
сооружениях, днищах судов и т.п. Этих гидробионтов называют бентосом. Бентос
делится на две большие группы - зообентос и фитобентос [Крылова, 1982]. Иногда
выделяют также протозойный и бактериальный бентос, биомасса которого в ряде
водоёмов достаточно велика, несмотря на малые размеры слагающих организмов
[Алимов, Финогенова, 1976].
Зообентос - это экологическая группа организмов,
характеризующихся связью с дном водных объектов как субстратом, на котором или
внутри которого организмы проводят свою жизнь. Организмы бентоса, пребывающие в
толще воды продолжительное время, относятся к группе планктонобентоса или
бентопланктона [Жадин, 1950].
Зообентосные организмы являются наиболее многочисленной
группой гидробионтов, имеющей огромное экологическое и хозяйственное значение.
Они потребляют формирующееся в водоёмах и приносимое извне органическое
вещество, осуществляют самоочищение водоёмов и водотоков, составляют основу
питания большинства видов рыб. Наконец, могут использоваться в пищу человеком,
на корм домашним животным и как промышленное сырьё [Свешников, 1989].
Традиционно зообентос разделяется на несколько групп в
зависимости от размеров - микробентос (менее 0,1 мм), мезобентос (0,1-2,0 мм) и
макробентос (более 2 мм) [Поляков, 1994].
В систематическом плане пресноводный макрозообентос состоит
из личинок амфибиотических насекомых (хирономиды и другие двукрылые, ручейники,
веснянки, стрекозы, подёнки, жесткокрылые), ракообразных, олигохет, моллюсков,
пиявок и некоторых других животных. По способу питания в нём встречаются
фитофаги, фильтраторы, хищники, некрофаги и сапрофаги, способные осуществлять
полный цикл круговорота веществ [Рылов, 1930].
Преобладающее большинство донных беспозвоночных животных
составляет группу так называемого "кормового зообентоса",
непосредственно используемого в пищу рыбами. Все эти деления в значительной
форме условны. Как показывали исследования последних лет, инфузории, другие
простейшие и даже микроорганизмы в составе детрита могут потребляться рыбами -
детритофагами. Максимальные размеры кормового бентоса определяются
максимальными размерами обитающих в водоёмах рыб - бентофагов [Арабина, 1988].
Бентос подразделяется на эпи - и инфауну. Представители
первой обитают в грунте, второй - внедряются, закапываются в него. Среди донных
форм часты колониальные виды, например, губки, многие кишечнополостные.
По приуроченности к определённому грунту различают пелофильные
формы, обитающие в иле, псаммофильные - живущие на песке, литофильные -
заселяющие камни и т.д. [Жадин, Герд, 1961].
Характер грунта, а также глубина, рельеф дна часто оказывают
определяющее влияние на состав и численность донных организмов.
Приспособления донных форм к жизни на грунте или в нём
сводятся к развитию средств, противостоящих сносу, действию волн, засыпанию
оседающей взвесью. Прикрепление к субстрату, вытягивания тела вверх,
заглубление в субстрат, уплощение тела, миграции донных животных - основные
способы их защиты от неблагоприятного воздействия среды [Арабина, 1988].
Прикрепление к субстрату характерно для простейших, губок,
некоторых иглокожих, моллюсков, кишечнополостных, водорослей и достигается
пневматическим способом, с помощью корневидных выростов, биссуса - клейкого
секрета. Оно бывает временным (актинии, двустворчатые моллюски) или постоянным
(асцидии) [Рылов, 1930].
Заглубление в субстрат отмечается у некоторых моллюсков,
иглокожих, червей личинок насекомых. Моллюски способны проникать в дерево или
камень на глубину до 0,5 м. Гидробионты, живущие на дне рек, приспосабливаются
к жизни на сильном течении за счёт крючьев, присосок, коготков [Крылова, 1982].
По отношению к субстрату в бентосе выделяют следующие группы:
- Прикреплённые организмы (сессильный
бентос). Обычно характерна вытянутая форма тела. Часто колониальные организмы,
как например, губки, мшанки, кораллы. Органы движения редуцируются или меняют
форму.
- Лежащие организмы. Отличаются сильно
расширенным и низким телом. К таким организмам относятся крабы, двустворчатые
моллюски и др.
- Закапывающиеся организмы. Черви,
голотурии, брюхоногие и двустворчатые моллюски, личинки насекомых и др.
Способность закапываться связана с морфологическими изменениями: сильная нога у
двустворчатых моллюсков и в связи с этим неплотно закрывающаяся раковина,
длинные сифоны, мощная мускулатура червей и др.
- Сверлящие организмы. В пресных водах
личинки некоторых насекомых минируют листья и стебли водных растений. Сверлящие
организмы по существу являются узниками своего жилища, в котором они
прокладывают ходы при помощи выделяемой кислоты или механически.
- Свободно двигающиеся организмы. Многие
животные передвигаются по дну при помощи различно устроенных конечностей
[Кузьмина, 2007].
Таким образом, на основе анализа имеющихся литературных
источников можно заключить, что зообентос - многочисленная и разнообразная
группа донных беспозвоночных, играющая важную роль как в морских, так и в
пресноводных экосистемах.
1.2
Таксономический состав и морфо-биологические особенности речного зообентоса
Основу речного макрозообентоса составляют личинки
амфибиотических насекомых, ракообразные, олигохеты, моллюски, пиявки и
некоторые другие животные. Рассмотрим основные группы речного зообентоса и их
морфологические и биологические особенности.
Малощетинковые черви или олигохеты (Oligochaeta).
Широко распространённая как в почве, так и на дне водоёмов
группа животных. Тело малощетинковых червей состоит из большого числа
сегментов. Впереди находятся предротовой и околоротовой сегменты, лишённые
щетинок. У зрелых особей имеется поясок, представляющий собой железистое
утолщение кожи. На каждом сегменте щетинки, как правило, в виде четырёх пучков
- двух спинных и двух брюшных [Шарова, 1999].
Олигохеты - донные животные, особенно многочисленные на
илистых грунтах. Наиболее характерной группой олигохет в водоёмах европейской
части страны являются представители семейства трубочники (Tubificidae).
Передним концом тела трубочники погружены в ил, а задний, служащий для дыхания,
выставляют над поверхностью дна, производя колебательные движения. Трубочники -
типичные грунтоеды. Погружаясь передним концом в грунт дна водоёма, они всё
время заглатывают ил с песком. Чем беднее грунт, тем больше его заглатывается
[Кутикова, 1977].
Обычно олигохеты наиболее многочисленны в водоёмах, сильно
загрязнённых органикой. В таких водоёмах высокой численности достигают Tubifex
tubifex, Limnodrilus udekemianus, Limnodrilus hoffmeisteri, относящиеся к
семейству Tubificidae и Lumbriсulus variegatus, являющийся
представителем семейства Lumbriсidae [Кузьмина, 2007].
Равноногие раки (Isopoda).
Внутреннее строение равноногих раков (или изопод) почти не
отличается от плана строения типичных высших раков. А к специфическим чертам
равноногих раков относятся поперечно-полосатая мускулатура, а также смещённое
назад сердце и эктодермальная кишка. Отсутствие карапакса у изопод обусловило
смещение сердце в заднюю часть переона, а дыхательная функция у них перенесена
на плеоподы. Кишка у них исключительно эктодермальная, и только
гепатопанкреатические железы эндодермальны [Шарова, 1999].
Представителем пресноводных равноногих ракообразных является
водяной ослик (Asellus aquaticus) - типичный обитатель большинства
стоячих и медленнотекущих водоёмов Европы [Смирнов, 1971].
Водяной ослик имеет уплощённую форму тела. Голова образована
слиянием всех головных сегментов с первым торакомером. Грудных сегментов семь.
Первые два брюшных сегмента заметны, остальные слиты с тельсоном. Антеннулы
короткие, антенны длинные. Четыре простых глаза расположены по бокам головы.
Грудных ног семь пар [Кутикова, 1977].
Водяные ослики - типичные растительноядные животные. Помимо
водных растений, они в большом количестве поедают листовой опад, падающий в
водоём [Смирнов, 1971].
Личинки подёнок (Ephemeroptera).
На территории России встречается около 200 видов подёнок
[Кузьмина, 2007].
У большинства видов подёнок личиночная жизнь продолжается
один год, но у некоторых, например Ephemera и Polymitarcys, - два
года.
Тело личинки чётко делится на подвижную голову, грудь и
брюшко. Для подёнок характерен гемиметаморфоз. Вышедшая из яйца личинка не
имеет трахейных жабр и даже зачатков крыльев. Имеется две латеральные хвостовые
нити. Дыхание осуществляется через покровы тела. После первой линьки появляются
зачатки трахейных жабр и срединная хвостовая нить. С каждой последующей линькой
происходят постепенные изменения в строение личинок. В течение личиночной жизни
проходит 20-25 линек [Шарова, 1999].
Большинство личинок питается водорослями и детритом. Длина
личинок подёнок обычно не превышает 10-15 мм, личинки родов Ephemera и
Polymitarcys достигают 20-30 мм [Рылов, 1930].
По образу жизни и строению подёнки делятся на несколько
групп.
- Личинки первой группы: живут в небольших
водоёмах, богатых растительностью, способны к быстрому плаванью. Обитают в
прудах, в небольших реках среди зарослей растений, за которые они держатся
своими длинными тонкими ногами. Среди плавающих личинок наиболее обычны
представители родов Cloeon, Leptophlebia.
- Личинки второй группы: ползающие,
приспособлены к жизни на илистом дне. Типичными представителями этой группы
являются личинки родов Ephemegella и Ordella. Они обитают в
озёрах и медленно текучих реках, преимущественно на илистых грунтах.
- Личинки третьей группа включают личинок,
прокладывающих ходы в глинистых берегах рек. Характерными представителями
являются личинки родов Ephemera, Palingenia. Почти всю свою жизнь роющие
личинки проводят в ходах, проделываемых ими в илистом грунте озёр и речек
[Кузьмина, 2007].
Личинки комаров (Nematocera).
Комаров относят к насекомым с полным превращением. Всё
развитие протекает в воде. Яйца откладываются поодиночке (Anopheles) или
склеенными в пакеты (Culex, Theobaldia) на поверхности воды или на
поверхности почвы на месте временных водоёмов. Рост личинок связан с тремя
линьками. Длительность личиночной жизни зависит от температурных условий, в
течение лета на севере бывает одна - две генерации, на юге - до шести и более
[Кузьмина, 2007].
Личинки комаров живут в самых разнообразных как стоячих и
слабо текущих водоёмах, так и в горных реках, некоторые виды выдерживают
загрязнение и значительное увеличение солёности воды [Шарова, 1999].
Тело личинки делится на голову, грудь и брюшко и несёт
многочисленные перистые волоски. По бокам головы расположены небольшие глазки и
хорошо развитые сложные глаза. Грудные сегменты у большинства личинок комаров
слиты между собой. Брюшко состоит из девяти сегментов, но восьмой и девятый
сегмент слиты между собой [Кутикова, Старобогатов, 1977].
Наиболее характерным компонентом зообентоса равнинных рек
являются личинки комаров из семейства хирономид (Chironomidae). Они
относятся к числу самых распространённых бентических организмов пресных вод.
Некоторые виды обитают в морях, многие выдерживают значительное загрязнение
воды.
Длина личинок хирономид колеблется от нескольких миллиметров
до 2-3 см. Голова сильно хитинизирована, резко отделена от других сегментов.
Грудных сегментов три, брюшных - 10. Личиночные глаза расположены по бокам
головы. Ротовое отверстие расположено на нижней стороне головы [Свешников,
Алигаджиев, 1989].
Большинство личинок окукливается в своих домиках. Куколки
хищных личинок свободно лежат на грунте или плавают в воде. Передний вздутый
отдел куколки образован головогрудью. На спинной стороне головогруди лежат
зачатки крыльев, загибающиеся на брюшке. Куколки хирономид поднимаются к
поверхности воды лишь ко времени вылупления крылатого насекомого [Шарова,
1999].
Личинки стрекоз (Odonata).
Стрекозы относятся к насекомым с неполным превращением. Яйца
откладываются непосредственно в воду или в ил. Сразу после откладки яиц
начинается развитие личинок. Количество личиночных стадий разное: от 10 до 14
линек. Продолжительность личиночной жизни от года до 2-3 лет [Кузьмина, 2007].
Личинки стрекоз - типичные пресноводные организмы. В
солоноватых водоёмах живут в виде исключения. Тело личинок подразделяется на
три части: голову, грудь и брюшко. Голова неподвижная, сросшаяся с грудью.
Глазки встречаются лишь у нескольких видов.
Грудь образована свободным первым и слившимися между собой
вторым и третьим сегментами. На груди располагаются зачатки крыльев. Брюшко
состоит из 11 сегментов [Кутикова, Старобогатов, 1977].
Водные жуки (Coleoptera).
Водные жуки обитают преимущественно в небольших стоячих и медленно
текучих эвтрофных водоёмах с богатой растительностью. Некоторые виды характерны
для быстротекучих рек и ручьёв.
Типичными представителями этой группы являются плавунцы -
водные жуки тёмно-бурого или оливкового цвета. Переднеспинка имеет жёсткие
края. Длина достигает 40 мм. Тело обтекаемой формы, уплощено в дорсовентральном
направлении. Сегменты тела надёжно соединены между собой. Большие фасеточные
глаза, расположенные по бокам головы. Грудь состоит из трёх сегментов. Крылья
первой пары образуют твёрдые надкрылья, а крылья второй пары предназначены для
полёта [Кузьмина, 2007].
Вертячки (Gyrinidae) плавают на поверхности воды, погружаясь
в воду при приближении опасности. Длина тела - 3,5-5,0 мм, форма обтекаемая.
Окраска чёрно-стального цвета [Яшнов, 1969].
Личинки водных жуков очень разнообразны по облику и строению.
Обычно они имеют ноги (иногда редуцированны), а по бокам или на конце брюшка
часто несут трахейные жабры [Кутикова, Старобогатов, 1977].
Водные клопы (Heteroptera).
В отряде преобладают наземные формы. Сравнительно небольшое
число видов клопов, например, представители семейства Hydrocorisae, обитают в
воде или на её поверхности (Amphibiocorisae - клопы-водомерки). Практически все
водные клопы и водомерки - хищники. Развитие всех фаз жизненного цикла проходит
в воде или на её поверхности. Водомерки зимуют на суше, остальные водные клопы
- в воде [Кутикова, Старобогатов, 1977].
Тело водных клопов уплощённое, тёмноокрашенное. На большой
голове находятся глаза и крепкий хоботок. Сильно развитые ноги передней пары
служат для схватывания добычи. Брюшко состоит из восьми сегментов. Крыльев -
две пары [Кутикова, Старобогатов, 1977].
Брюхоногие моллюски (Gastropoda).
Брюхоногие моллюски характеризуются огромным разнообразием
видов, занимая самые разнообразные биотопы. Это асимметричные животные с ясным
разделением тела на голову, ногу и внутренностный мешок, убранный внутрь
раковины. Раковина спирально завитая или колпачковидная [Кутикова,
Старобогатов, 1977].
Двустворчатые моллюски (Bivalvia).
Насчитывается около 15 тысяч видов двустворчатых. Большое
морфологическое разнообразие связано с различными условиями обитания, глубиной
и с качеством воды.
Наиболее обычными представителями пресноводных двустворчатых
моллюсков являются беззубки (Anodonta) и перловицы (Unio) [Яшнов,
1969].
1.3
Особенности развития зообентоса степных рек
Характерными организмами для зообентоса степных рек являются
олигохеты, личинки хирономид, подёнки, моллюски. Это связано с тем, что в реках
степной зоны уменьшается скорость течения водотоков, возрастает зарастаемость
макрофитами, что приводит к снижению в них доли организмов реофильного
комплекса [Белюченко, Мамась, 2005].
В зообентосе степных рек преобладают типичные фито - и
пелофильные виды, а из других групп - двукрылые, моллюски, представители
лимнофильного бентоса - жуки и клопы [Алимов, 1976].
При продвижении от истока рек к устью отмечается сокращение
численности типичных реобионтов, обитающих на твёрдых субстратах и преобладание
видов, характерных для малопроточных участков рек, встречающихся в прибрежных
зарослях водоёмов, на заиленных биотопах [Баканов, 1996].
Динамику количественных показателей зообентоса малых рек
определяют, в основном, особенности развития хирономид и олигохет,
преобладающих в реках. Доля хирономид, олигохет, нематод, моллюсков в бентосе
равнинных рек степной зоны возрастает.
На распределение зообентоса оказывают влияние скорость
течения, биотопическая неоднородность, заиленность зарегулированных участков
рек, зарастаемость макрофитами, изменение гидрохимических параметров. Характер
формирования структуры доминирующего комплекса бентосных организмов также
формируется в условиях изменения физико-химических параметров в летний период и
зарегулирования рек в устьевом участке, что отражается на общем снижении
количественного развития бентоса и распределении донных организмов [Яковлев,
1988].
На формирование видовой структуры бентоса оказывают влияние
притоки, малая проточность, однообразие биотопов и зарегулированность рек
временными земляными плотинами.
Степные реки характеризуется значительными антропогенными
нагрузками с сельскохозяйственных полей, зарастаемостью растительностью -
камышом, тростником, рогозом, осокой. Бентофауна в связи с этим имеет
значительную численность личинок хирономид и олигохет. Создаются оптимальные
условия для развития лимнофильной донной фауны. В силу этого преимущественное
развитие получают пелофильные организмы. Высокой численности достигают широко
распространённые эврибионтные виды олигохет и хирономид, способные развиваться на
разнообразных биотопах [Белюченко, 2005].
В условиях антропогенного воздействия на речную систему
пространственная направленность и интенсивность изменений донных сообществ
меняются в зависимости от силы, времени и характера воздействия. На всем
протяжении реки доминантами являются эвритопные виды, обитатели эвтрофных
водоёмов. На отдельных участках развиваются сообщества с олигодоминатным
характером популяционных распределений.
Реки степной зоны характеризуются сравнительно однородными
условиями, при которых процессы преобразования русла не выражены, что приводит
к преобладанию лимнофильной фауны. Там, где реки имеют относительно высокие
скорости течения, формируется обилие специфичной фауны [Алимов, 1976].
Зарегулирование водотоков ведёт к перестройке типичных речных
биоценозов и процветанию фауны, характерной для антропогенно-трансформированных
водоёмов, что негативно сказывается на многих реофильных видах и ведёт к
унификации бентофауны рек [Белюченко, 2005].
Проведённые в 2010-2014 гг. гидробиологические съёмки
показали, что зообентос в степных реках формируется, в основном, за счёт
личинок насекомых и крупных двустворчатых моллюсков.
В реках Бейсуг, Ея, Кирпили, Челбас практически на всех
участках отмечались личинки хирономид: Tendipes plumosus, Tendipes
dorsalis, виды, обитающие обычно в грунтах, богатых органикой, питающиеся
фильтрационным способом органическими взвесями и детритом и играющие большую
роль в процессе самоочищения воды [Москул, 2013].
Кроме личинок хирономид, в исследуемых водоёмах встречались
личинки подёнок, стрекоз, мошек, настоящих комаров и прочих двукрылых
насекомых. В местах со стоячей водой, на грунтах, богатых перепревшей
растительностью, в массе развиваются малощетинковые черви (Olygochaeta) родов Limnodrilus
и Tubifex, что характерно для эвтрофных водоёмов. В верхнем течении рек
Ея, Кирпили, Челбас единично встречались личинки ручейников, представителей
семейства Limnophilidae. На дне заиленных участков рек Челбас и Кирпили
отмечали крупных двустворчатых моллюсков: Unio ovalis, Unio
longirostris, Anodonta sp., дающих большую биомассу общего бентоса.
Большое развитие во всех водоёмах получил моллюск Dreissena polymorpha,
особенно в предустьевых участках рек Ея, Челбас [Болкунов, Рудакова, Ерзиков,
2014].
Благодаря исследованиям сотрудников Кубанского
государственного университета и Краснодарского филиала ФГУП "ВНИРО",
были выполнены работы, позволяющая определить численность и биомассу зообентоса
рек Азово-Кубанской равнины.
Они установили, что донная фауна рек Азово-Кубанской равнины
представлена в основном малощетинковыми червями (Oligochaeta) и хирономидами
(Chironomidae), а также личинками водных жуков, стрекоз и моллюсками.
Остаточная биомасса кормового зообентоса (Oligochaeta, Chironomidae и др.)
исследуемых водоёмов колеблется от 1,9±0,23 г/м2 (р. Албаши) до
3,4±0,25 г/м2 (р. Кирпили), составляя в среднем 2,6±0,28 г/м2 или
26±0,30 кг/га (таблица 1) [Рыбохозяйственное освоение., 2013].
Таблица 1 - Численность и биомасса зообентоса рек
Азово-Кубанской равнины [Рыбохозяйственное освоение., 2013]
|
Река (количество обработанных проб)
|
Численность, экз. /м2
|
Биомасса
|
|
|
г/м2
|
кг/га
|
|

|
|
|
|
Ея (n=62)
|
389,5±1,24
|
2,6±0,24
|
26±0,22
|
|
Челбас (n=62)
|
456,7±1,12
|
3,1±0,37
|
31±0.34
|
|
Бейсуг (n=68)
|
524,8±1,62
|
2,9±0,31
|
29±0,28
|
|
Кирпили (n=68)
|
516,3±0,98
|
3,4±0,25
|
34±0,24
|
|
Понура (n=45)
|
324,5±1,15
|
2,2±0,36
|
22±0,36
|
|
Албаши (n=34)
|
298,8±1,32
|
1,9±0,23
|
19±0,22
|
|
Ясени (n=34)
|
372,6±1,21
|
2,1±0,31
|
21±0,28
|
|
Средняя
|
411,88±0,96
|
2,6±0,28
|
26±0,30
|
Донная фауна степных рек Краснодарского края представлена как
мягкими, так и жёсткими организмами. Остаточная биомасса мягкого зообентоса
(хирономиды и олигохеты) колебалась в пределах от 0,34 до 3,05 г/мІ и в среднем
составила 2,38 г/м2 [Болкунов, Рудакова, Ерзиков, 2014].
1.4
Антропогенное влияние на зообентос степных рек
Зообентос - один из основных индикаторов последствий не
только хозяйственной деятельности, но и климатических изменений, современные и
ожидаемые масштабы которых значительны. Поэтому оценка закономерностей и причин
динамики зообентоса актуальны как в научном, так и практическом отношении
[Водные ресурсы, 2011].
На современном этапе степные реки Краснодарского края, в том
числе и Бейсуг, испытывают одни и те же воздействия: они загрязняются бытовыми
и промышленными водами, стоками с сельскохозяйственных угодий; неправильными
вырубками нарушаются водоохранные зоны; водные ресурсы используются не
рационально [Борисов, 1971; Белюченко, 2005].
Анализ сложившейся ситуации позволяет из множества негативных
процессов и явлений, порожденных и обусловленных человеческой деятельностью в
степной части Краснодарского края, выделить и назвать главные равнозначные по
актуальности группы: изменение речного стока, обмеление и заиление русла,
зарастание русла, подтопление прилегающих территорий, повышенная минерализация
и химическое загрязнение речной воды, эвтрофикация водоёмов [Райнин, 2000]. Все
выше перечисленные проблемы характерны и для бассейна реки Бейсуг, территории
которого характеризуется высоким процентом антропогенной преобразованности
[Фильчагов, 1989].
Проблема истощения речного стока очень актуальна в настоящее
время для степных рек. Половодье в них наблюдается в весенний период, а в
засушливое время года некоторые участки рек пересыхают и образуют мелководные
разобщённые плёсы, которые интенсивно зарастают тростником, рогозом, осоками и
водорослями. Причём процесс обмеления продолжается, общий объем стока
уменьшается, и летнее пересыхание становится всё более частым и
продолжительным. Это объясняется тем, что питание степных рек в основном происходит
за счёт атмосферных осадков и грунтовых вод [Коровин, 1990].
Ранее степные реки текли по территории, обильно покрытой
травянистой и древесной растительностью, но с интенсивным освоением территории
Кубанских степей естественная растительность была уничтожена. Густой травостой
предохранял реки от заиления, прирусловая древесная растительность регулировала
речной сток и защищала водоёмы от прямых солнечных лучей [Борисов, 1973;
Данилов, 1983; Маккавеев, 1988; Маккавеев, Чалов, 1988].
Немаловажным фактором давления на речные системы является
разработка в пойме реки карьеров по добыче строительного и иного материала и
его вывоз, что ведёт к нарушению ландшафтов поймы на больших площадях.
Уничтожение естественной растительности и распашка земель в
водоохранных зонах привело к увеличению степени водной и ветровой эрозии.
Развитие эрозионных процессов в значительной степени зависит и от
сельскохозяйственных культур, возделываемых на склонах [Гайдай, 2005]. Смыву
почвы на склонах способствуют обработка земель, в результате которой пахотный
горизонт разрыхляется, наличие плохо проницаемого нижнего горизонта, а также
слабая защита почвы растениями [Белюченко, 2005].
Эта проблема очень актуальна, так как распахивание водосборов
привело к сокращению водного стока и увеличению водной и ветровой эрозии почв
[Барышман, 1983]. В этих условиях транспортирующаяся способность степных рек
оказалась недостаточной, что привело к отложению части твёрдого стока и наносов
в руслах рек. Процессу заиления способствовало увеличение площади водотоков
путём строительства дамб. Слой ила в реках достигает 1,5-5,2 м, что является
одной из основных проблем вторичного загрязнения речной воды. При существующей
зарегулированности степных рек вынос твёрдого стока в их устья доходит до 5 %
[Белюченко, Мамась, 2005].
В бассейне реки Бейсуг в последнее столетие не отмечаются
наводнения. Причиной этого стало зарегулирование стока и высокая заиленность
рек бассейна. Конец 50-х и начало 60-х гг. ХХ в. характеризовалось усилением
зарегулированности стоков практически всех рек степной зоны края и замедлением
их течения на большом протяжении, что обусловило заболачивание значительных
территорий и подтоплением многих долинных площадей [Нежиховский, 1989].
Зарегулированность естественного стока привела к тому, что
степные реки плотинами поделили на короткие участки (всего их на реке Бейсуг
насчитывается более 300), в которых изменились естественные гидрологические
условия, снизилась пропускная способность. Процессы накопления донных отложений
во вновь образованных прудах стали преобладающими, что привело к сильному
заилению и обмелению русла реки. В настоящее время большинство прудов в
бассейне реки представляют собой мелководные (средние глубины 0,8-1,5 м)
заболачиваемые водоёмы, многие из которых используются для разведения товарной
рыбы [Абаев, 1992].
Обмеление рек ведёт к более интенсивной деградации их русел.
Помимо накопления ила, фарватеры рек зарастают тростником и камышом, что еще
более затрудняет их сток. В таких условиях когда-то богатые ихтиофауной степные
реки практически утратили своё природное и хозяйственное значение. Вода рек
теряет своё качество, становится непригодной как для целей воспроизводства рыб,
так и для водоснабжения и орошения [Кузнецов, Шевченко, Кадышев, 2000].
Угроза антропогенного эвтрофирования водоёмов стала
осознаваться только во второй половине прошлого столетия. Для водоёмов,
особенно с малой проточностью, избыточное поступление биогенных веществ не
менее опасно, чем токсическое загрязнение. Когда содержание в воде фосфора, азота,
калия превышает критический уровень, ускоряются жизненные процессы водных
организмов. Как следствие, получают массовое развитие планктонные водоросли
("цветение" воды), вода приобретает неприятный запах и привкус, её
прозрачность снижается, увеличивается цветность, повышается содержание
растворенных и взвешенных органических веществ. Перенасыщение воды
органическими соединениями стимулирует развитие сапрофитных бактерий (в том
числе особо опасных - болезнетворных), водных грибов, резко обостряя эпидемиологическую
обстановку на водных объектах [Водные ресурсы, 2004].
Потепление вод рек вызывает усиление плотностей стратификации
вод, уменьшение концентраций кислорода в придонных слоях и, в конечном счёте,
некоторое понижение биомасс ракообразных, червей и моллюсков.
Большие иловые отложения в руслах рек закрыли существующие
выходы грунтовых вод. Количество поступающих грунтовых вод в воды реки
снижается, отчего происходит их обмеление [Горелов, 1961].
Характерной особенностью большинства гидрофобных органических
веществ и тяжёлых металлов, поступающих в водные объекты, является способность
их аккумулироваться в донных отложениях в результате физического и химического
фракционирования; седиментации взвешенных частиц, сорбирующих их растворенные
формы; сорбции на поверхности донных отложений и т.д. [Гайдай, 2006].
Как правило, накопление веществ происходит в донных
отложениях водоёмов с замедленным течением, заиленных участках, дельтах и
эстуариях. Загрязнение донными отложениями является источником передачи тяжёлых
металлов и гидрофобных загрязняющих веществ (пестицидов, полициклических
ароматических углеводородов, нефтепродуктов и др.) по трофической цепи: ил -
растения - беспозвоночные - мирные рыбы - хищные рыбы. Концентрации веществ в
каждом последующем звене цепи могут возрастать в 10 и более раз. Являясь средой
обитания бентосных организмов, такие донные отложения могут непосредственно
влиять на их видовой состав и разнообразие [Крылова, 1982].
Проблема изучения накопления загрязняющих веществ в донных
отложениях становится особо острой в настоящее время, когда в результате
интенсивной антропогенной деятельности человека большинство донных отложений
водных объектов оказалось загрязнёнными. Химический состав органических
загрязняющих веществ и тяжёлых металлов в донных отложениях довольно чётко
отражает специфику источников загрязнения водных объектов и более наглядно по
сравнению с водными массами характеризует их экологическое состояние. Последнее
в значительной мере обусловлено тем, что в пресноводных объектах по сравнению с
морскими объектами осаждение загрязняющих веществ преобладает над
трансформацией их состава в процессе седиментации [Водные ресурсы, 2007].
Вода степных рек по химическому составу отличается
значительной щёлочностью (рН до 8,4), что обуславливает её высокую жёсткость. В
воде степных рек много нитратов (от 44 до 68 мг/л), что связано со сносом в их
русла азота и органики с окружающих полей, где периодически вносятся удобрения.
Следует особо подчеркнуть, что вода рек от устья до истока характеризуется
высокими значениями ХПК (от 24 до 41 мг/л) и БПК5 (от 13 до 23
мг/л). Содержание фенолов в воде устьевой части рек нередко превышает ПДК.
Содержание некоторых тяжелых металлов в воде (например, марганец) превышает
уровень ПДК, но по большинству элементов их уровень ниже ПДК. Вода содержит
большое количество взвешенных веществ на всем протяжении рек. Источниками
поступления органических веществ в воду являются бытовые и животноводческие
стоки и т.д. [Волошина, 2007].
Речная вода сильно загрязнена органикой (численность
сапротрофных микроорганизмов составляет 107 КОЕ/мл) и относится к
полисапробной [Волошина, 2006].
Донные отложения содержат большие количества
аммонифицирующих, амилолитических и сульфатредуцирующих бактерий. Наиболее
широко встречаются представители родов Pseudomonas, Bacillus, Flavobacterium.
Значительное количество сульфатредуцирующих бактерий свидетельствует о
накоплении органического вещества, что ведёт к интенсивному восстановлению
сульфата и выделения сероводорода и является негативным фактором развития
систем [Чалов, Чернов, 1994].
2.
Физико-географическая характеристика реки Бейсуг
В нижнем плейстоцене (примерно 1,2 млн лет назад) широкая
долина реки Кубань уже отделяла северную Прикубанскую равнину от южной и, тем
самым, отрезала её от влияния геологических процессов, происходящих в горах
Кавказа. На Прикубанской равнине в конце нижнего плейстоцена заложилась
самостоятельная речная сеть (Бейсуг, Челбас, Ея и др.) [Чередниченко, 1988].
Приустьевая зона бассейна реки, как в голоцене, так и на
современном этапе переживает инверсию движений. На месте вала заложен
Бейсугский лиман, зона плавней и разливов Челбаса, Бейсуга и других степных
рек, которым соответствуют современные опускания до 1-2 мм/год; в сторону
Тимашевска, структурная ступень испытывает современные поднятия до 1,5 мм/год
[Лилиенберг, 1979].
В нижней части бассейна реки (плавневая зона) выделяется
рельеф, созданный озёрно-лиманной аккумуляцией, представленный заболоченной
равниной. Его формирование происходило в условиях преобладающих новейших
погружений. На поверхности выделяются озёрно-лиманные формы рельефа,
представленные существующими и зарастающими лиманами и озёрами и слабо
выпуклыми межлиманными возвышенностями. В целом плавневый район характеризуется
плоскоравнинным рельефом [Чупахин, 1974]. Совершенно не имеющие уклона, плоские
недренированные лиманные заболоченности имеют отметки поверхности 0,2-0,9 м.
Межлиманные возвышенности (гряды) представляют собой узкие 50-100 м
полого-выпуклые незатопляемые участки продолговатой или неправильной формы
[Борисов, 1978].
Река Бейсуг - третья по длине река Азово-Кубанской равнины,
является типичной стенной рекой, имеющей свыше 20 крупных и малых притоков, и
относится к бассейну Азовского моря. Она берет начало в 9 км на северо-запад от
города Кропоткина и, проделав путь длиной 243 км, впадает в районе станицы
Бриньковской в Бейсугский лиман, который соединяется с Ясенским заливом
Азовского моря. Река имеет два крупных притока первого порядка: Левый Бейсужёк,
Правый Бейсужёк, а также ряд малых притоков и балок [Мельникова, 2003]. В
бассейне реки свыше 300 прудов, суммарная площадь их зеркальной поверхности
около 0,2 тыс. км2, а суммарный объём воды - около 128 тыс. м3
[Нагалевский, 1992; Ресурсы поверх, вод., 1973]. Бейсугский лиман образовался в
результате затопления морем устьевого пространства реки Бейсуг. От Азовского
моря он отделён Ясенской косой [Борсов, 1978].
Бассейн реки Бейсуг целиком расположен в пределах
Краснодарского края и по своей конфигурации напоминает овал, повёрнутый длинной
осью с востока на запад (рисунок 1).
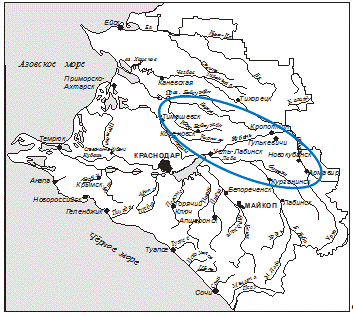
Рисунок 1 - Гидрологическая сеть Краснодарского края и
бассейн реки Бейсуг (выделен овалом)
Длина бассейна, считая по прямой линии, составляет 153 км, а
наибольшая ширина - 61 км. Площадь водосборного бассейна - около 5190 км2.
Границами бассейна служат: на юге - бассейн реки Кирпили, на севере - бассейн
реки Челбас. Водораздельные пространства имеют высоту от 60 до 150 м [Симов,
1989].
Берега реки в основном пологие, высотой не более 5-7 м в
верхнем течении, 7-8 м в среднем течении и 5-6 м в нижнем течении. В нижней
части бассейна реки много плавней, что объясняется широкой долиной, а также
выходом реки из берегов (вследствие паводков из-за сильных дождей), а также в
связи с тем, что узкое Бейсугское гирло не успевает пропустить всю воду в
Бейсугский лиман [Белюченко, 2005].
В геоморфологическом отношении территория бассейна реки
Бейсуг находится в центральной части Азово-Кубанской низменности,
представляющей собой волнистую платообразную поверхность, наклонённую к западу
и северо-западу, прорезанную долинами её притоков, сухих балок и оврагов,
образующих отдельные локальные водоразделы. На поверхности равнины отмечается
сильная эрозионная деятельность, плоскостной смыв и просадочные явления
[Горелов, 1961].
Оврагообразование на склонах не отмечается, а образование
небольших промоин в период ливней и снеготаяния отмечается как на крутых
склонах, так и на откосах земляных плотин. Водной эрозии берегов в силу зарегулированности
рек и отсутствия интенсивного течения не отмечается. Волновая переработка
берегов незначительна из-за зарастания берегов и большей части прудов болотной
растительностью.
Процессы подтопления земель и заболачивания пониженных
участков в бассейне реки Бейсуг получили широкое распространение. Наибольшее
развитие они имеют на поверхности современной, верхнечетвертичной и
среднечетвертичной дельты. Основная причина подтопления - слабая естественная
дренированность территории. Дополнительное влияние на подъем уровня грунтовых
вод оказало строительство прудов, сильное заиливание русла, орошение земель,
потери воды из водопроводов и другие факторы, связанные с антропогенной
деятельностью. В условиях небольшой водопроводимости грунтовых вод и увеличения
инфильтрационного питания в два раза наблюдается подъем уровня грунтовых вод на
2,5-6,0 м [Кондратьев, Попов, Снищенко, 1982]. Вся территория бассейна реки
расположена в средней части Краснодарского края и входит в состав Кавказского,
Тбилисского, Выселковского, Кореновского, Тимашевского, Брюховецкого,
Каневского и Приморско-Ахтарского административных районов. Большая часть
площади бассейна полностью утратила свой естественный природный ландшафт в
результате интенсивного хозяйственного использования. Около 2/3 территории
бассейна занято сельскохозяйственными угодьями, большая часть которых
представляет собой пахотные земли. Бассейн реки Бейсуг достаточно густо
заселён, а большинство населённых пунктов располагается вдоль берегов реки.
Благодаря своим климатическим и почвенным условиям основная его площадь занята
под сельскохозяйственное производство [Вагинов, 1976].
Сельскохозяйственное использование почв пойм, балок и оврагов
зачастую осуществляется без изучения закономерностей развития их рельефа,
гидрологии, глубины почвенного покрова и уровня залегания грунтовых вод. Важным
аспектом давления на вышеперечисленные территории является строительство дамб,
дорог, животноводческих комплексов, распашка балок и решение других
хозяйственных нужд [Белюченко, 2004].
Транспортная сеть рассматриваемого региона представлена
разветвлённой сетью автомобильных и железных дорог, трубопроводов. Через
бассейн проходят оживлённые железнодорожные магистрали, автодороги федерального
значения "Дон", Краснодар - Ростов-на-Дону с интенсивным движением
автотранспорта. Большая интенсивность движения предполагает большие количества
загрязнений, которые, попадают во все компоненты речного бассейна [Вальков,
1996; Дегтярева, 1991].
Водный режим отличается значительными колебаниями уровней и
расходов воды в течение года. Половодье чаще всего начинается в феврале, за
счёт таяния снежного покрова, нередко сопровождается выпадением жидких осадков.
Ввиду неустойчивости зим обычно бывает несколько волн половодья. В результате
этого половодье сравнительно невысокое и значительно растянуто по времени (в
среднем 2,5 месяца). Окончание половодья в среднем приходится на конец апреля
начало мая. Сток половодья в сильной степени зарегулирован каскадом прудов.
Продолжительность основной волны половодья обычно составляет 15-30 суток.
Первые ледяные образования на реке Бейсуг и его притоках отмечаются во второй
половине ноября, которые, как правило, разрушаются через 10-15 дней. Во второй
половине декабря происходит повторное образование ледостава, в конце февраля
начале марта - третье. Средняя дата вскрытия реки - середина марта [Коровин,
1981; Ресурсы поверхностных вод., 1973].
Основными причинами летней межени являются высокая
температура воздуха и малое количество атмосферных осадков, а также заборы воды
на орошение, водоснабжение и заполнение прудов [Коровин, 1981; Ресурсы
поверхностных вод., 1973].
Глубина залегания грунтовых вод изменяется в пределах 0,1-2,0
м. Питание грунтовых вод исключительно инфильтрационное. Разгрузка их
осуществляется за счёт дренажа реками и балками, испарения, транспирации и
перетекания в напорные водоносные горизонты.
Бассейн реки Бейсуг, за исключением его западной плавневой
части, лежит в полосе степей.
Ихтиофауна реки довольно скудная, так как гидрологический и
гидрохимический режим водотока весьма неблагоприятен. При протяжённости реки
всего 243 км на ней насчитывается 301 запруда. Многочисленные запруды
превратили реки в каскад застойных прудов с нарушенной системой самоочищения от
бытовых и хозяйственных стоков [Плотников, 2001].
Строительство дамб на реке Бейсуг привело к тому, что она
практически перестала выполнять роль в воспроизводстве рыб Азовского моря.
Также практически полностью была уничтожена береговая растительность, а берега
распахиваются до уреза воды. Все эти антропогенные вмешательства привели к
заилению и обмелению реки. Применение химикатов на прилегающих
сельскохозяйственных угодьях, размещение животноводческих комплексов на
территории водосбора, а также сброс неочищенных сточных вод в реку Бейсуг привели
к сильному загрязнению [Водные ресурсы, 2007].
Формирование ихтиофауны рек Северо-Западного Кавказа
протекало за счёт рыб, проникших сюда в третичном периоде из водоёмов
Восточно-Европейской равнины. Современная ихтиофауна водоёмов бассейна по
существующему зоогеографическому делению континентальных водоёмов относится к
Дунайско-Кубанскому участку Черноморского округа [Берг, 1962].
По данным П.С. Матвиенко [1961], в середине 1950-х годов в
реке Бейсуг общее число видов и подвидов рыб составляло 31. В настоящее время в
реке обитает 16 видов без учёта интродуцированных рыб: щука, плотва,
краснопёрка, линь, пескарь, уклейка, густера, лещ, верховка, карась золотой и
серебряный, сазан, щиповка, окунь, судак. В низовьях реки ихтиофауна намного
разнообразнее за счёт видов, заходящих из лимана.
3. Материал и
методы исследования
Материалом для данной выпускной квалификационной работы
послужили пробы зообентоса степной реки Бейсуг (рисунок 2), отобранные в летний
и осенний периоды 2014 г.
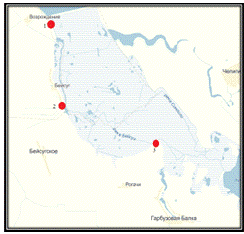
- нижнее течение р. Бейсуг; 2 - среднее течение р. Бейсуг; 3
- верхнее течение р. Бейсуг
Рисунок 2 - Места отбора проб зообентоса в реке Бейсуг
Пробы для данного исследования отбирали в летний период -
02.07.14 г. и 04.08.14 г., и в осенний период - 06.09.14 г. Мероприятия по
отбору проб проводили в вечерние часы суток в нескольких точках, примерно
соответствующих нижнему, среднему и верхнему течению реки (рисунок 3, таблица
2).
Таблица 2 - Характеристика станций отбора проб
|
№ п/п
|
Местоположение станций
|
Глубина отбора проб, м
|
Характер грунта
|
Характеристика станций
|
|
1
|
Нижнее течение в районе хутора Возрождение
|
0,25-0,40
|
песчано - илистый
|
Вода зеленовато-жёлтая, Малое наполнение русла,
средняя глубина - 1,55 м
|
|
2
|
Среднее течение в районе хутора Бейсуг
|
1,00-1,45
|
глиняно - илистый
|
Вода зеленовато-жёлтая, Среднее наполнение
русла, средняя глубина - 0,75 м
|
|
3.
|
Верхнее течение в районе посёлка Гарбузовая
Балка
|
0,65-1,45
|
заиленный песок
|
Вода зеленовато-жёлтая, Среднее наполнение
русла, средняя глубина - 0,80 м
|
Для отлова беспозвоночных использовался
гидробиологический скребок. Диаметр скребка - 0,45 м, длина металлической ручки
- 1,2 м. В каждой точке отбора проб с его помощью зачерпывали донные отложения
с бентосными организмами.
Отбор проб производили на расстоянии
0,5-3,0 м от берега и на глубине 0,1-1,5 м. Также на каждом участке обследовали
камни и зону уреза воды, погруженные в воду древесные сучья и стволы, листья
водных растений.
Для выборки фауны из грунта пробу перед промывкой подвергали
отмучиванию. После этого остаток грунта в тазу просматривали, всех оставшихся
животных выбирали, а грунт выбрасывали.
После отбора животных помещали в пластмассовую ёмкость и
фиксировали 4 % формалином из расчёта одна часть 40 % формалина на девять
частей воды. Пробу снабжали этикеткой, на которой обозначали время, дату и
место отбора пробы, температуру воды и глубину места взятия пробы.
Сведения о каждой пробе записывали в журнал исследований.
Таким образом, сбор первичного материала осуществлялся в соответствии с
методиками, общепринятыми в гидробиологии [Абакумова, 1983; Методические
рекомендации по изучению., 1989].
При лабораторной обработке проб использовали методику,
описанную в работе В.И. Жадина [1960]. Перед разборкой количественных проб
формалин аккуратно сливался через газ, а проба заливалась водой. Фракцию в
ёмкости перед разборкой взбалтывали и наливали тонким слоем в чашку Петри.
Разборка проводилась под микроскопом МБС: под увеличением в 8 и 16 раз.
Животных выбирали тонким хирургическим пинцетом и распределяли по видам или
группам. В каждой группе просчитывали общее число животных. После этого
полученные данные суммировались и, тем самым, определяли численность всех
организмов в пробе. Затем производится перерасчёт на один квадратный метр
площади дна реки.
Биомасса отдельных групп бентоса определялась взвешиванием на
электронных весах после обсушивания на фильтровальной бумаге до исчезновения
влажных пятен на ней. Общая биомасса организмов, обнаруженных в пробе,
определялась суммированием и пересчитывалась на 1 м2 площади дна.
Всю полученную информацию заносили в таблицы для дальнейшей обработки.
Расчёт численности зообентосных организмов на 1 м2 обловленной
площади реки проводили по формуле (1):
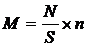 , (1)
, (1)
где М - численность организмов в 1 м2;
N -
количество организмов в пробе (экземпляров);
n -
количество взятых скребков;
S - площадь
обловленной территории (м2), находимая по формуле (2):
 ,
,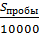 (2)
(2)
где S - площадь обловленной территории (м2);
Sпробы - площадь пробы (см2);
- площадь дна в 1 м2, выраженная в см2.
Подсчёт биомассы зообентоса на 1 м2 обловленной
площади реки (г/м2) производили по формуле (3):
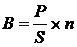 , (3)
, (3)
где B - биомасса организмов в 1 м2;
P - масса
организмов (г);
S - площадь
обловленной территории (м2);
n -
количество взятых скребков.
Определение организмов зообентоса проводилось с помощью
соответствующих определителей зообентоса [Панкратова, 1977; Кирейчук, 2001;
Полоскин, Хаитов, 2006].
Собранные материалы обработаны стандартными методами вариационной
статистики [Лакин, 1973].
Для количественной оценки сложности структуры сообществ
бентических животных определялись индексы Шеннона и Маргалефа [Одум, 1986].
Коэффициент сходства систематического состава сообщества
определялся по формуле Сёренсена-Чекановского (4):
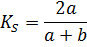 (4)
(4)
где Ks - коэффициент Сёренсена-Чекановского; - число
видов в одном сообществе;- число видов в другом сообществе.
Пределы этого коэффициента от 0 до 1, причём Ks =
1 означает полное сходство сообществ (абсолютное совпадение списков), а Ks
= 0 означает, что они ни имеют ни одного общего вида.
4.
Современное состояние зообентоса реки Бейсуг и его значение в питании массовых
видов рыб
4.1 Видовой
состав зообентоса
Таксономический состав исследованных нами проб зообентоса из
р. Бейсуг был следующим:
тип кольчатые черви - Annelida
класс пиявки - Hirudinea
отряд хоботные пиявки - Rhynchobdellida
семейство хоботные пиявки - Rhynchobdellea
пиявка улитковая - Glossiphonia complanata
тип моллюски - Mollusca
класс Брюхоногие - Gastropoda
отряд лёгочные улитки - Panpulmonata
семейство катушки - Planorboidae
катушка роговая - Planorbarius corneus
семейство прудовики - Lymnaeidae
обыкновенный прудовик - Limnaea stagnalis
класс двустворчатые моллюски - Bivalvia
отряд униониде - Unionida
семейство перловицы, беззубки - Unionidae
перловица - Unio pictorum
тип членистоногие - Arthropoda
класс ракообразные - Crustaceae
отряд разноногие раки - Amphipoda
семейство гаммариды - Gammaridae
бокоплав - Gammarus lacustris
класс паукообразные - Arachnoidae
отряд клещи - Acarina
семейство водяные клещи - Hydrachnidae
водяной клещ - Limnochares aquatica
класс насекомые - Insecta
отряд жесткокрылые - Coleoptera
семейство водолюбы - Hydrophilidae
малый водолюб - Hydrophilus sp.
семейство плавунцы - Dytiscidae
плавунец окаймлённый - Dytiscus marginalis
отряд клопы - Hemiptera
семейство гладыши - Nolonectidae
гладыш обыкновенный - Notonecta glauca
отряд стрекозы - Odonata
семейство коромысла - Aeshnidae
синее коромысло - Aeschna cyanea
отряд двукрылые - Diptera
семейство комары-звонцы - Chironomidae
звонец мохнатоусый - Chironomus plumosus
хирономус семиредуктус - Chironomus semireductus
Таким образом, за период исследований в реке было обнаружено
12 видов зообентосных организмов, относящихся к трём типам, шести классам и
девяти отрядам. Наибольшим видовым разнообразием характеризовались насекомые,
представленные четырьмя отрядами и пятью видами.
Качественный состав исследованных нами проб зообентоса в
разных участках р. Бейсуг представлен в таблице 3.
Повсеместно (100 % проб) встречались личинки хирономид
Chironomus plumosus, и лишь в шести пробах из девяти (67 %) были отмечены
личинки хирономид Chironomus semireductus. В семи пробах из девяти
(встречаемость - 78 %) были обнаружены катушка роговая и перловица. В
трёх пробах (33 %) отмечены бокоплавы, плавунцы, в двух пробах (22 %) -
обыкновенный прудовик, улитковая пиявка, малый водолюб, личинки стрекоз.
Водяные клещи и клопы были обнаружены лишь в одной пробе из девяти (11 %).
Таблица 3 - Видовой состав зообентоса разных участков
продольного профиля реки Бейсуг в летне-осенний период 2014 года
|
Вид
|
Участок реки
|
|
нижнее течение
|
среднее течение
|
верхнее течение
|
|
класс пиявки - Hirudinea
|
|
отряд хоботные пиявки - Rhynchobdellida
|
|
Glossiphonia complanata - пиявка улитковая
|
+
|
-
|
-
|
|
класс брюхоногие - Gastropoda
|
|
отряд лёгочные улитки - Panpulmonata
|
|
Planorbarius corneus - катушка роговая
|
+
|
+
|
+
|
|
Limnaea stagnalis - прудовик обыкновенный
|
+
|
+
|
-
|
|
класс двустворчатые моллюски - Bivalvia
|
|
отряд униониде - Unionida
|
|
Unio pictorum - перловица
|
+
|
+
|
+
|
|
отряд клещи - Acarina
|
|
Limnochares aquatica - водяной клещ
|
-
|
+
|
-
|
|
класс ракообразные - Crustaceae
|
|
отряд разноногие раки - Amphipoda
|
|
Gammarus lacustris - бокоплав
|
+
|
+
|
-
|
|
класс насекомые - Insecta
|
|
отряд двукрылые - Diptera
|
|
Chironomus plumosus - звонец мохнатоусый
|
+
|
+
|
+
|
|
Chironomus semireductus - хирономус семиредуктус
|
-
|
+
|
+
|
|
отряд клопы - Hemiptera
|
|
Notonecta glauca - гладыш обыкновенный
|
+
|
-
|
-
|
|
отряд стрекозы - Odonata
|
|
Aeschna cyanea - синее коромысло
|
+
|
-
|
-
|
|
отряд жесткокрылые - Coleoptera
|
|
Hydrophilus caraboides - водолюб малый
|
+
|
+
|
-
|
|
Dytiscus marginalis - плавунец окаймлённый
|
+
|
+
|
-
|
|
Всего
|
10
|
9
|
4
|
Установлено, что видовое разнообразие зообентоса сильно
отличалось по участкам реки: в нижнем течении отмечено 10 видов, в среднем -
девять, а в верхнем - только четыре.
В составе зообентоса нижнего течения р. Бейсуг отмечены
клопы, пиявки и личинки стрекоз, не представленные в районах среднего и
верхнего течения. В то же время, водяные клещи были обнаружены только в среднем
течении.
Более высокое видовое разнообразие зообентоса в нижнем и
среднем течении р. Бейсуг можно объяснить тем, что в указанных участках река
образует широкие плёсы и заливы, имеет большее разнообразие биотопов (заросли
макрофитов на различных грунтах, песчаные и илисто-песчаные субстраты без
макрофитов и др.), лучшую прогреваемость, что способствует большему видовому
разнообразию организмов зообентоса.
В верхнем течении преобладает биотоп илистых грунтов, который
предпочитают в основном личинки хирономид. Для многих других видов зообентосных
организмов такой биотоп непригоден [Жадин, 1950].
В таблице 4 представлены результаты подсчёта значений
коэффициента Сёренсена, с помощью которого была оценена степень видового
сходства зообентоса между отдельными участками течения реки.
Таблица 4 - Значения коэффициента Сёренсена при сравнении
видового состава зообентоса верхнего, среднего и нижнего течения реки Бейсуг
|
Сравниваемые участки
|
Величина индекса
|
|
Верхнее - среднее течение
|
0,62
|
|
Верхнее - нижнее течение
|
0,43
|
|
Среднее - нижнее течение
|
0,74
|
Коэффициент сходства Сёренсена - бинарная мера сходства,
предложенная Торвальдом Сёренсеном в 1948 году. Это безразмерный показатель,
применяемый в биологии для количественного определения степени сходства
биологических объектов. Также известен под названиями: мера ассоциации, мера
подобия и др. Чем выше его величина, тем сходство между сравниваемыми объектами
больше [Дулепов, 2004].
Как показано в таблице 4, при анализе полученных значений
коэффициента Сёренсена было выявлено, что наибольшее сходство видового состава
зообентоса характерно для среднего и нижнего участков продольного профиля реки,
а наименьшее - для верхнего и нижнего. Эти результаты позволяют заключить, что
среднее и нижнее течение реки Бейсуг более близки по гидрологическим
характеристикам, а верхнее течение занимает несколько обособленное положение.
При этом в верхнем течении встречались только те виды,
которые имели широкое распространение и были отмечены во всех участках
продольного профиля реки (таблица 3).
Видовой состав зообентоса в реке имел также определённую
сезонную динамику, что демонстрируют данные таблицы 5. Летний и осенний периоды
характеризовались различным количеством видов в зообентосе.
В нижнем участке течения реки наблюдалось резкое снижение
видового разнообразия от лета к осени с девяти до трёх видов организмов. В
верхнем участке снижение было менее значительным: с четырёх видов до трёх. В
среднем участке течения реки видовое разнообразие по сезонам не изменялось.
Таблица 5 - Сезонная динамика видового состава зообентоса
реки Бейсуг в летне-осенний период 2014 года
|
Вид
|
Встречаемость по участкам реки и сезонам
|
|
нижнее течение
|
среднее течение
|
верхнее течение
|
|
лето
|
осень
|
лето
|
осень
|
лето
|
осень
|
|
Пиявка улитковая - Glossiphonia complanata
|
+
|
-
|
-
|
-
|
-
|
-
|
|
Катушка роговая - Planorbarius corneus
|
+
|
-
|
+
|
+
|
+
|
-
|
|
Обыкновенный прудовик - Limnaea stagnalis
|
+
|
-
|
+
|
+
|
-
|
-
|
|
Перловица - Unio pictorum
|
-
|
+
|
+
|
-
|
+
|
+
|
|
Бокоплав - Gammarus lacustris
|
+
|
-
|
+
|
-
|
-
|
-
|
|
Водяной клещ - Limnochares aquatica
|
-
|
-
|
-
|
+
|
-
|
-
|
|
Малый водолюб - Hydrophilus caraboide
|
+
|
-
|
+
|
-
|
-
|
-
|
|
Плавунец окаймлённый - Dytiscus marginalis
|
+
|
+
|
-
|
+
|
-
|
-
|
|
Гладыш обыкновенный - Notonecta glauca
|
+
|
-
|
-
|
-
|
-
|
-
|
|
Синее коромысло - Aeschna cyanea
|
+
|
-
|
-
|
-
|
-
|
-
|
|
Звонец мохнатоусый - Chironomus plumosus
|
+
|
+
|
+
|
+
|
+
|
+
|
|
Хирономус семиредуктус - Chironomus
semireductus
|
-
|
-
|
+
|
+
|
+
|
+
|
|
Всего
|
9
|
3
|
7
|
6
|
4
|
3
|
Пять видов зообентоса были отмечены в реке только в летний
период. В эту группу входили пиявка улитковая, бокоплавы, малый водолюб, гладыш
обыкновенный и синее коромысло (таблица 5). Этот факт был связан либо с
сезонностью развития организмов (только в наиболее тёплое летнее время года),
либо с их редкостью в реке.
Водяные клещи отмечены только в осенний период. Остальные
шесть видов зообентоса сезонной динамики встречаемости не имели (таблица 5).
4.2
Пространственная динамика количественных характеристик зообентоса реки Бейсуг
Установлено, что количественные показатели развития
зообентоса в реке зависели от её участка и отличались в верхнем, среднем и
нижнем течении. Наиболее высокие количественные характеристики зообентоса были
отмечены в верхнем и нижнем участках реки (рисунок 3). Среднее количество
донных беспозвоночных на этих участках было выше по сравнению со средним
течением на 54,2 и 41,7 экз. /м2 соответственно.
На наш взгляд, отличия в численности зообентоса на разных участках
продольного профиля реки объясняются следующими факторами.
Нижнее течение достаточно мелководно, вода в этом участке
реки хорошо прогревается, здесь имеются разнообразные биотопы с обильным
развитием водных растений (рогоз, камыш, элодея и др.), создающих
дополнительные места обитания.
Теплолюбивые виды, к которым относятся все летне-осенние
бентические организмы, предпочитают менее глубоководные участки реки для
развития, где имеются более благоприятные условия для развития и роста личинок,
а также большее количество кормовых организмов.
Всё это и приводит к высокой численности зообентосных
организмов в нижнем течении водотока.
В верхнем течении р. Бейсуг причины высокой численности
зообентоса другие. Этот участок водотока характеризуется преобладанием илистых
субстратов и низким уровнем видового разнообразия зообентоса (всего четыре
вида), что приводит к снижению межвидовой конкуренции и увеличению численности
отдельных видов. Такая закономерность известна для многих зообентосных
сообществ, особенно находящихся под антропогенным прессом [Безматерных, 2007].

Рисунок 3 - Пространственная динамика численности зообентоса
в разных участках продольного профиля реки Бейсуг
В таблице 6 приведены результаты сравнения средних значений
численности зообентоса в трёх исследованных участках реки Бейсуг с помощью
t-критерия Стъюдента.
Результаты сравнения не выявили статистически достоверных
отличий в количественных показателях развития зообентоса в верхнем, среднем и
нижнем течении реки. Этот факт связан, вероятно, с малым количеством отобранных
проб в сочетании с высокой изменчивостью численности организмов в отдельных
пробах.
Таблица 6 - Оценка достоверности отличий в средних значениях
численности зообентоса в разных участках реки Бейсуг с помощью t-критерия
Стъюдента
|
Участок реки
|
Нижнее течение
|
Среднее течение
|
Верхнее течение
|
|
Нижнее течение
|
-
|
1,05
|
0,17
|
|
Среднее течение
|
1,05
|
-
|
0,76
|
|
Верхнее течение
|
0,17
|
0,76
|
-
|
|
Примечание: tст = 1,96 для уровня
значимости 5 % и tст = 2,58 для уровня значимости 1 %
|
В таблице 7 приведены данные о численности отдельных видов
зообентоса в разных точках отбора проб (нижнее, среднее и верхнее течение). Как
видно из приведённой таблицы, отдельные виды характеризовались различной
численностью в зависимости от участка реки.
Таблица 7 - Численность зообентоса в разных участках
продольного профиля реки Бейсуг в летне-осенний период в экз. /м2
|
Численность
|
Численность по участкам реки
|
|
нижнее течение
|
среднее течение
|
верхнее течение
|
|
Пиявка улитковая
|
12,5
|
0,0
|
0,0
|
|
Катушка роговая
|
8,3
|
20,8
|
12,5
|
|
Обыкновенный прудовик
|
8,3
|
16,7
|
0,0
|
|
Перловица
|
12,5
|
16,7
|
29,2
|
|
Бокоплав
|
29,2
|
4,2
|
0,0
|
|
Водяной клещ
|
0,0
|
4,2
|
0,0
|
|
Малый водолюб
|
4,2
|
8,3
|
0,0
|
|
Плавунец окаймлённый
|
16,7
|
8,3
|
0,0
|
|
Гладыш обыкновенный
|
8,3
|
0,0
|
0,0
|
|
Синее коромысло
|
16,7
|
0,0
|
0,0
|
|
Звонец мохнатоусый
|
33,3
|
21,6
|
80,8
|
|
Хирономус семиредуктус
|
0,0
|
7,5
|
40,0
|
|
Итого
|
150,0
|
108,3
|
162,5
|
Такие виды, как звонец мохнатоусый, пиявка улитковая,
бокоплав, плавунец окаймлённый, гладыш обыкновенный и синее коромысло были
наиболее многочисленными в нижнем течении. Катушка роговая, прудовик
обыкновенный, водяной клещ и малый водолюб были наиболее многочисленны в
среднем течении. Перловица и два вида хирономид - звонец мохнатоусый и
хирономус семиредуктус максимальных показателей численности достигали в верхнем
течении реки (таблица 7).
Таким образом, в разных участках реки сформировались
зообентосные сообщества, которые характеризовались разным количественным
соотношением видов. Структуру сообществ можно описать, определив относительную
численность отдельных видов, т.е. выразив её в процентах.
Данные по структуре зообентосных сообществ в отдельных
участках реки Бейсуг представлены в таблице 8.
Таблица 8 - Соотношение отдельных видов зообентоса по
численности в разных участках продольного профиля реки Бейсуг в процентах
|
Вид
|
Численность по участкам реки
|
|
нижнее течение
|
среднее течение
|
верхнее течение
|
|
Пиявка улитковая
|
8,3
|
0,0
|
0,0
|
|
Катушка роговая
|
5,6
|
19,2
|
7,7
|
|
Обыкновенный прудовик
|
5,6
|
15,4
|
0,0
|
|
Перловица
|
8,3
|
15,4
|
18,0
|
|
Бокоплав
|
19,4
|
3,9
|
0,0
|
|
Водяной клещ
|
0,0
|
3,9
|
0,0
|
|
Малый водолюб
|
2,8
|
7,7
|
0,0
|
|
Плавунец окаймлённый
|
11,1
|
7,7
|
0,0
|
|
Гладыш обыкновенный
|
5,6
|
0,0
|
0,0
|
|
Синее коромысло
|
11,1
|
0,0
|
0,0
|
|
Звонец мохнатоусый
|
22,2
|
19,9
|
49,7
|
|
Хирономус семиредуктус
|
0,0
|
6,9
|
24,6
|
|
Итого
|
100,0
|
100,0
|
100,0
|
В нижнем течении реки по численности преобладали
(доминировали) два вида: представитель семейства хирономид - звонец мохнатоусый
и бокоплав. Субдоминирующее положение занимали плавунец окаймлённый и синее
коромысло (таблица 8).
В среднем течении реки доминирующее положение в составе
зообентоса по численности звонца мохнатоусого сохранилось. А второй
доминирующий вид сменился. Место бокоплава заняла катушка роговая. Субдоминирующее
положение также занимали моллюски - обыкновенный прудовик и перловица (таблица
8).
Зообентосное сообщество верхнего течения реки носило
хирономидный характер. Доминирующим по численности видом также являлся звонец
мохнатоусый. На долю личинок этого вида пришлось почти 50 % от общего
количества организмов зообентоса. Субдоминирующим по количеству видом являлся
хирономус семиредуктус (таблица 8).
Таким образом, характерной особенностью структуры
зообентосного сообщества р. Бейсуг по численности являлось преобладание личинок
хирономид и брюхоногих моллюсков. По всем участкам реки руководящую роль по
численности играл являлся звонец мохнатоусый.
4.3
Пространственная динамика биомассы зообентоса реки Бейсуг
Установлено, что значения биомассы зообентоса в реке зависели
от её участка и отличались в верхнем, среднем и нижнем течении. Наиболее
высокие значения биомассы зообентоса были отмечены в верхнем и среднем участках
реки (рисунок 4). Среднее значение биомассы беспозвоночных на этих участках
было выше по сравнению с нижним течением на 135,97 и 72,69 г/м2
соответственно.
На рисунке 4 показаны среднее значение биомассы зообентоса в
разных участках реки Бейсуг в период исследований.
Высокие значения биомассы зообентоса в среднем течении р.
Бейсуг можно объяснить тем, что в указанных участках река образует широкие
плёсы и заливы, вода в этом участке реки хорошо прогревается, здесь имеется
большое разнообразие биотопов. Всё это и приводит к высокому значению биомассы
зообентоса в данных участках водотока.
На дне данных участков реки отмечали брюхоногих и крупных
двустворчатых моллюсков, насекомых, дающих большую биомассу.
Бентос заиленных грунтов верхнего течения р. Бейсуг
характеризуется высокой биомассой, так как представлен в основном
двустворчатыми моллюсками и личинками хирономид, дающими большую биомассу
общего бентоса. Для многих других видов зообентосных организмов такой биотоп
непригоден для обитания.

Рисунок 4 - Среднее значение биомассы всего зообентоса в
разных участках реки Бейсуг в период исследований
В таблице 9 приведены результаты сравнения средних значений
биомассы зообентоса в трёх исследованных участках реки Бейсуг с помощью
t-критерия Стъюдента.
Результаты сравнения не выявили статистически достоверных
отличий в показателях биомассы зообентоса в верхнем, среднем и нижнем течении
реки. Этот факт связан, вероятно, с малым количеством отобранных проб в
сочетании с высокой изменчивостью биомассы организмов в отдельных пробах.
Таблица 9 - Оценка достоверностей отличий в средних значениях
биомассы зообентоса в разных участках реки Бейсуг с помощью t-критерия
Стъюдента
|
Участок реки
|
Нижнее течение
|
Среднее течение
|
Верхнее течение
|
|
Нижнее течение
|
-
|
0,25
|
0,58
|
|
Среднее течение
|
0,25
|
-
|
0,26
|
|
Верхнее течение
|
0,58
|
0,26
|
-
|
|
Примечание: tст = 1,96 для уровня
значимости 5 % и tст = 2,58 для уровня значимости 1 %
|
На дне заиленных участков реки были отмечены крупные
двустворчатые моллюски Unio pictorum (жёсткий зообентос). Этот вид,
обитает обычно в грунтах, богатых органикой, питается фильтрационным способом
органическими взвесями, детритом, играет большую роль в процессе самоочищения
воды и даёт большую биомассу общего бентоса, но не является кормом для рыб.
На рисунке 5 показаны среднее значение биомассы кормового
зообентоса в разных участках реки Бейсуг в период исследований.
Высокие значения биомассы кормового или мягкого зообентоса в
нижнем и среднем течении р. Бейсуг можно объяснить тем, что на дне данных
участков реки отмечали крупных брюхоногих моллюсков и насекомых, дающих большую
биомассу общего бентоса.
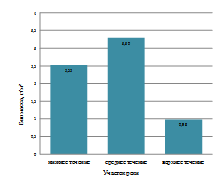
Рисунок 5 - Среднее значение биомассы кормового зообентоса в
разных участках реки Бейсуг в период исследований
В таблице 10 приведены данные о биомассе отдельных видов
зообентоса в трёх исследуемых участках реки Бейсуг. Как видно из приведенной
таблицы, биомасса зообентоса меняется в зависимости от исследованного участка
реки.
В нижнем течении р. Бейсуг наибольшую биомассу имели такие
виды, как пиявка улитковая, бокоплав, плавунец окаймлённый, гладыш
обыкновенный, обыкновенный прудовик и катушка роговая. Катушка роговая,
прудовик обыкновенный, плавунец окаймлённый имели наибольшую биомассу в среднем
течении. Два вида хирономид - звонец мохнатоусый и хирономус семиредуктус
максимальных показателей биомассы достигали в верхнем течении реки (таблица
10).
Таблица 10 - Биомасса различных групп зообентоса продольного
профиля реки Бейсуг в летне-осенний период в г/м2
|
Численность
|
Район отбора проб
|
|
нижнее течение
|
среднее течение
|
верхнее течение
|
|
Пиявка улитковая
|
0, 208
|
0,000
|
0,000
|
|
Катушка роговая
|
0,117
|
0,338
|
0, 208
|
|
Обыкновенный прудовик
|
1,458
|
2,708
|
0,000
|
|
Перловица
|
145,833
|
208,333
|
283,333
|
|
Бокоплав
|
0,188
|
0,042
|
0,000
|
|
Водяной клещ
|
0,000
|
0,013
|
0,000
|
|
Малый водолюб
|
0,050
|
0,033
|
0,000
|
|
Плавунец окаймлённый
|
0,270
|
0,137
|
0,000
|
|
Гладыш обыкновенный
|
0,133
|
0,000
|
0,000
|
|
Синее коромысло
|
0,025
|
0,000
|
0,000
|
|
Звонец мохнатоусый
|
0,069
|
0,019
|
0,402
|
|
Хирономус семиредуктус
|
0,000
|
0,010
|
0,373
|
|
Всего
|
148,351
|
211,633
|
284,316
|
Таким образом, в реке сформировались зообентосные сообщества,
которые характеризовались разной биомассой организмов по отдельным участкам
реки. Структуру сообществ можно описать, определив относительную биомассу
отдельных видов, т.е. выразив её в процентах.
Данные по биомассе зообентосных сообществ в отдельных
участках реки Бейсуг представлены в таблице 11.
Таблица 11 - Соотношение отдельных видов зообентоса по
биомассе в разных участках продольного профиля реки Бейсуг в процентах
|
Вид
|
Биомасса по участкам реки
|
|
нижнее течение
|
среднее течение
|
верхнее течение
|
|
Пиявка улитковая
|
0,14
|
0,00
|
0,00
|
|
Катушка роговая
|
0,08
|
0,16
|
0,08
|
|
Обыкновенный прудовик
|
0,98
|
1,27
|
0,00
|
|
Перловица
|
98,30
|
98,44
|
99,65
|
|
Бокоплав
|
0,13
|
0,02
|
0,00
|
|
Водяной клещ
|
0,00
|
0,01
|
0,00
|
|
Малый водолюб
|
0,03
|
0,02
|
0,00
|
|
Плавунец окаймлённый
|
0,18
|
0,06
|
0,00
|
|
Гладыш обыкновенный
|
0,09
|
0,00
|
0,00
|
|
Синее коромысло
|
0,02
|
0,00
|
0,00
|
|
Звонец мохнатоусый
|
0,05
|
0,01
|
0,14
|
|
Хирономус семиредуктус
|
0,00
|
0,01
|
0,13
|
|
Итого
|
100,00
|
100,00
|
100,00
|
В нижнем течении реки по биомассе преобладала (доминировала)
перловица. Субдоминирующее положение занимали обыкновенный прудовик и пиявка
улитковая (таблица 11).
В среднем течении реки доминирующее положение в составе
зообентоса по биомассе перловицы сохранилось. А второй доминирующий вид
сменился. Место пиявки улитковой заняла катушка роговая. Субдоминирующее
положение также занимал обыкновенный прудовик (таблица 11).
Зообентосное сообщество верхнего течения реки по биомассе
носило "моллюсковый" характер. Доминирующим по биомассе видом
являлась перловица. На долю организмов этого вида пришлось 98,44 % от общего
количества организмов зообентоса. Субдоминирующим по биомассе видом являлись
личинки хирономид - звонец мохнатоусый, хирономус семиредуктус (таблица 11).
Таким образом, характерной особенностью структуры
зообентосного сообщества р. Бейсуг по биомассе являлось преобладание
двустворчатых моллюсков. По всем участкам реки преобладающей по биомассе
являлась перловица.
4.4 Сезонная
динамика развития зообентоса реки Бейсуг
Рисунок 6 отражает сезонные изменения численности зообентоса
исследованного района в течение всего периода наших исследований.
Согласно представленному графику, в районе нижнего течения
реки количество донных беспозвоночных к осени постепенно уменьшалось.
Численность организмов в июле была выше по сравнению с августом на 62,5 экз. /м2,
а в августе по сравнению с сентябрём на 75,0 экз. /м2. Снижение
численности зообентоса осенью в сравнении с летним периодом связанно с
исчезновением в пробах пиявок, ракообразных и личинок стрекоз.
В районе среднего течения реки была отмечена обратная
тенденция: количество донных беспозвоночных к осени постепенно увеличивалось.
Численность организмов в августе по сравнению с июлем была выше на 25,0 экз. /м2,
а в сентябре по сравнению с августом на 50,0 экз. /м2. Возрастание
численности зообентоса осенью в сравнении с летним периодом связанно с
увеличением количества насекомых, брюхоногих и двустворчатых моллюск.
В районе верхнего течения р. Бейсуг наибольшее количество
донных беспозвоночных было обнаружено в июле. Численность организмов в этом
месяце была выше по сравнению с августом и сентябрём на 187,5 экз. /м2
соответственно. Снижение численности зообентоса осенью в сравнении с летним
периодом связанно с снижением количества брюхоногих моллюск и личинок хирономид
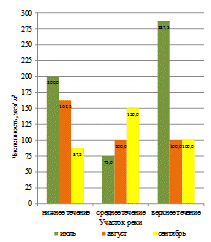
Рисунок 6 - Динамика количественных показателей зообентоса в
разные месяцы
В таблице 12 представлена сезонная динамика количественных
показателей отдельных видов зообентоса в разных участках реки Бейсуг.
Как видно из таблицы 12, наиболее долгий период развития
зообентоса характерен для личинок хирономид и моллюсков. Их развитие происходит
в период с июля по сентябрь. Пиявки и гладыш обыкновенный достигали наибольшей
численности в июле.
Таблица 12 - Сезонная динамика чисенности отдельных видов
зообентоса реки Бейсуг в экз. /м2
|
Вид
|
Нижнее течение
|
Среднее течение
|
Верхнее течение
|
|
VII
|
VIII
|
IX
|
VII
|
VIII
|
IX
|
VII
|
VIII
|
IX
|
|
Пиявка улитковая
|
25,0
|
12,5
|
0,0
|
0,0
|
0,0
|
0,0
|
0,0
|
0,0
|
0,0
|
|
Катушка роговая
|
0,0
|
25,0
|
0,0
|
12,5
|
12,5
|
37,5
|
25,0
|
12,5
|
0,0
|
|
Обыкновенный прудовик
|
25,0
|
0,0
|
0,0
|
0,0
|
50,0
|
0,0
|
0,0
|
0,0
|
0,0
|
|
Перловица
|
0,0
|
0,0
|
37,5
|
0,0
|
0,0
|
50,0
|
0,0
|
25,0
|
62,5
|
|
Бокоплав
|
50,0
|
37,5
|
0,0
|
12,5
|
0,0
|
0,0
|
0,0
|
0,0
|
0,0
|
|
Водяной клещ
|
0,0
|
0,0
|
0,0
|
0,0
|
0,0
|
12,5
|
0,0
|
0,0
|
0,0
|
|
Малый водолюб
|
0,0
|
12,5
|
0,0
|
0,0
|
25,0
|
0,0
|
0,0
|
0,0
|
0,0
|
|
Плавунец окаймлённый
|
25,0
|
0,0
|
25,0
|
0,0
|
0,0
|
25,0
|
0,0
|
0,0
|
0,0
|
|
Гладыш обыкновенный
|
25,0
|
0,0
|
0,0
|
0,0
|
0,0
|
0,0
|
0,0
|
0,0
|
0,0
|
|
Синее коромысло
|
25,0
|
25,0
|
0,0
|
0,0
|
0,0
|
0,0
|
0,0
|
0,0
|
0,0
|
|
Звонец мохнатоусый
|
25,0
|
50,0
|
25,0
|
32,0
|
9,2
|
18,5
|
210,0
|
10,0
|
30,0
|
|
Хирономус семиредуктус
|
0,0
|
0,0
|
0,0
|
18,0
|
3,3
|
6,5
|
52,5
|
12,5
|
7,5
|
|
Всего
|
200,0
|
162,5
|
87,5
|
75,0
|
100,0
|
150,0
|
287,0
|
60
|
100,0
|
Перловицы начинали выявляться в составе зообентоса реки с
августа, а наибольшей численности достигали в сентябре. Пик развития бокоплавов
приходился на июль. Малый водолюб в наибольшем количестве отмечен в августе.
Личинки хирономид демонстрировали чёткое снижение численности от июля к
сентябрю. У остальных видов определённых закономерностей в сезонных изменениях
количественных показателей не выявлено (таблица 12).
На рисунке 7 изображён график относительного количества
отдельных таксономических групп зообентоса в районе нижнего течения реки в
разные месяцы.
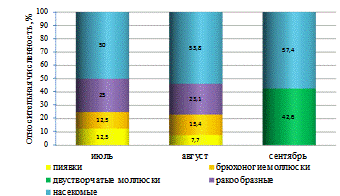
Рисунок 7 - Динамика относительной численности донных
организмов в составе зообентоса нижнего течения реки Бейсуг
Как показано на графике, в июле в данном районе по численности
доминировали насекомые (50,0 %). Вторыми по численности были ракообразные. Они
составляли 25,0 % от общего количества зообентоса в пробе. Остальные организмы
были представлены одинаковым количеством (12,5 %), кроме двустворчатых
моллюсков и паукообразных, которые отсутствовали.
В августе доминирующими видами в данном районе были также
насекомые - 53,8 %, второе место по количеству организмов в пробе занимали
ракообразные - 23,1 %, на третьем месте по численности находились брюхоногие
моллюски - 15,4 %, пиявки - 7,7 %, а двустворчатые моллюски и паукообразные
также отсутствовали.
В сентябрьской пробе появились раковины двустворчатых
моллюск, которые составили 42,6 %. Насекомые были представлены в количестве
57,4 %, остальные представители зообентоса исчезли.
На рисунке 8 показан график относительного количества
отдельных групп зообентоса в районе среднего течения реки Бейсуг в разные
месяцы.
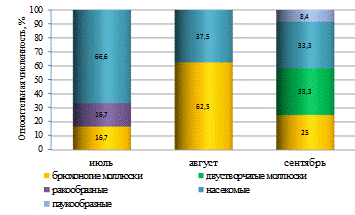
Рисунок 8 - Динамика относительной численности донных
организмов в составе зообентоса среднего течения реки Бейсуг
В июле также как и в районе нижнего течения преобладали
насекомые (66,6 %), ракообразные и брюхоногие моллюски имели одинаковое
количество и составляли по 16,7 % от общего количество зообентоса в пробе.
Остальные представители отсутствовали.
В составе августовской пробы доминировали брюхоногие моллюски
(62,5 %), субдоминантами по численности были насекомые.
В сентябре 33,3 % всех организмов приходилось на долю
насекомых. Двустворчатые моллюски также составляли 33,3 %. На третьем месте по
численности - брюхоногие моллюски (25,0 %), паукообразные составляли 8,4 %.
На рисунке 9 изображен график относительного количества
отдельных таксономических групп зообентоса в районе верхнего течения реки.
В июле так же, как и в нижнем и среднем течение,
превалировали насекомые (91,3 %), на втором месте по численности были
брюхоногие моллюски.
В августе доминирующими видами также были насекомые - 62,5 %.
Субдоминирующими по численности были двустворчатые моллюски (25 %), брюхоногие
составили 12,5 %.
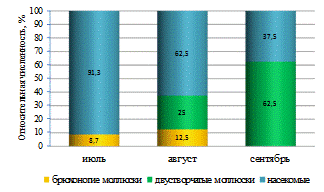
Рисунок 9 - Динамика относительной численности донных
организмов в составе зообентоса в районе верхнего течения
В сентябре 62,5 % всех организмов приходилось на долю
двустворчатых моллюск. Насекомые составили 37,5 %.
В таблице 13 представлена сезонная динамика биомассы
различных групп зообентоса в разных участках продольного профиля реки Бейсуг в
летне-осенний период.
Как видно из таблицы 13, наибольшая биомасса зообентоса
характерна для двустворчатых моллюсков. Их развитие происходит в период с
августа по сентябрь. Плавунец окаймлённый и катушка роговая достигали
наибольшей биомассы в сентябре. Гладыш обыкновенный выявлялся в составе
зообентоса реки только в июле и достигал при этом высокой биомассы. Пик
увеличения биомассы обыкновенного прудовика приходился на август.
Бокоплавы наибольшую биомассу достигали в августе. Личинки
хирономид демонстрировали чёткое снижение биомассы от июля к сентябрю. У
остальных видов определённых закономерностей в сезонных изменениях биомассы не
выявлено (таблица 13).
На рисунке 10 изображён график динамики соотношения биомассы
отдельных таксономических групп зообентоса в районе нижнего течения реки в
разные месяцы.
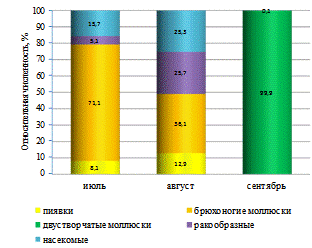
Рисунок 10 - Динамика относительной биомассы донных
организмов в составе зообентоса нижнего течения реки Бейсуг
Как показано на графике, в июле в данном районе по биомассе
доминировали брюхоногие моллюски (71,1 %). Вторыми по биомассе были насекомые.
Они составляли 15,7 % от общей биомассы зообентоса в пробе. Пиявки и
ракообразные составляли 8,1 % и 5,1 % от общей биомассы организмов 12,5 %.
Двустворчатые моллюски и паукообразных отсутствовали.
В августе доминирующими видами по биомассе в данном районе
были также брюхоногие моллюски - 36,1 %, второе место по биомассе организмов в
пробе занимали ракообразные и насекомые - 25,7 % и 25,3 %, на третьем месте по
численности находились пиявки - 12,9 %. Двустворчатые моллюски и паукообразные
также отсутствовали.
В сентябрьской пробе появились раковины двустворчатых
моллюск, биомасса которых составляла 99,9 %. Насекомые составляли 0,1 %
биомассы, остальные представители зообентоса исчезли.
На рисунке 11 показан график динамики биомассы отдельных
групп зообентоса в районе среднего течения реки Бейсуг в разные месяцы.
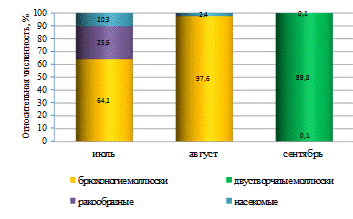
Рисунок 11 - Относительная биомасса донных организмов в
составе зообентоса в районе среднего течения
В июле также как и в районе нижнего течения по биомассе
преобладали брюхоногие моллюски (64,1 %), биомасса ракообразных составляла 25,6
%, а насекомых 10,3 % от общей биомассы зообентоса. Остальные представители
отсутствовали.
В составе августовской пробы по биомассе доминировали
брюхоногие моллюски (97,6 %), субдоминантами были насекомые.
В сентябре 99,8 % биомассы всех организмов приходилось на
долю двустворчатых моллюск. Насекомые и брюхоногие моллюски составляли по 0,1 %
биомассы, биомасса паукообразных была незначительна.
На рисунке 12 изображен график относительной биомассы отдельных
таксономических групп зообентоса в районе верхнего течения реки в период
исследований.
В июле в данном районе по биомассе превалировали насекомые
(84,6 %), на втором месте по численности были брюхоногие моллюски.
В августе доминирующее положение по биомассе заняли
двустворчатые моллюски - 99,8 %. Субдоминирующими по численности были насекомые
(0,1 %) и брюхоногие (0,1 %).
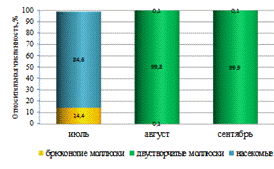
Рисунок 12 - Относительная биомасса донных организмов в
составе зообентоса в районе верхнего течения
В сентябре также 99,9 % по биомассе всех организмов
приходилось на долю двустворчатых моллюск. Насекомые составили 0,1 %.
Таким образом, в сезонной динамике развития зообентосного
сообщества р. Бейсуг прослеживаются следующие тенденции:
) снижение численности бентических организмов к осени, за
счёт уменьшения количества амфибиотических насекомых, которые покидают водоём;
) наблюдается увеличение биомассы бентических организмов, за
счёт интенсивного роста моллюск.
4.5 Оценка
экологического состояния зообентоса реки Бейсуг
Для оценки современного состояния зообентоса р. Бейсуг на
первом этапе работ были определены такие показатели, как число видов, индекс
Шеннона и индекс Маргалефа [Одум, 1986]. С помощью указанных показателей было
проведено сравнение трёх участков реки (таблица 13).
Таблица 13 - Основные показатели состояния зообентоса р.
Бейсуг в разных участках продольного профиля
|
Показатель
|
Участок реки
|
|
нижнее течение
|
среднее течение
|
верхнее течение
|
|
Число видов
|
10
|
9
|
4
|
|
Индекс Шеннона
|
0,94
|
0,92
|
0,50
|
|
Индекс Маргалефа
|
0,42
|
0,44
|
0,14
|
Установлено, что число видов, а также величины обоих
указанных выше индексов оказались максимальными в нижнем и среднем течении
реки. В верхнем течении величины этих показателей были минимальными.
Полученные данные можно объяснить следующим образом.
Индекс Маргалефа предложен, исходя из положения, что с
увеличением уровня загрязнения водоема видовое богатство, как правило, падает.
Индекс Маргалефа принимает максимальное значение, если все особи принадлежат к
разным видам и минимальное, когда все особи принадлежат к одному виду [Одум,
1986].
Индекс Шеннона суммирует большое количество информации о
численности и видовом составе организмов, учитывая число видов и степень их
доминирования. Возрастание величины индекса указывает на увеличение
неопределенности и однородности структуры изучаемой системы, наоборот, снижение
его величины показывает, что структура становится менее однородной и возрастает
доминирование каких-то ее отдельных элементов. В первом случае мы имеем дело с
более сложной системой [Одум, 1986].
Следовательно, можно заключить, что в наиболее благополучном
состоянии находятся зообентосные сообщества нижнего и среднего течения реки.
Это можно объяснить тем, что в указанных участках, река имеет песчаные и илисто-песчаные
грунты, являющиеся более благоприятными биотопами для жизни бентических
организмов.
В верхнем течении преобладают сильно заиленные грунты,
которые для многих зообентосных организмов непригодены для жизни. Пойма
верхнего течения реки окружена большим количеством сельскохозяйственных угодий,
с стоками которых попадает большое количество пестицидов и ядохимикатов, что
усиливает заиление и зарастание русла, повышенная минерализация и химическое
загрязнение речной воды [Райнин, 2000].
Для сравнения показателей развития зообентоса в р. Бейсуг с
другими реками региона мы сравнили полученные нами результаты с данными
сотрудников Кубанского государственного университета и Краснодарского филиала
ФГУП "ВНИРО", которые были получены в 2003-2007 годах на различных
водотоках Азово-Кубанской равнины (таблица 14).
Проведённые нами исследования показали, что за данный
промежуток времени количество зообентосных организмов в р. Бейсуг уменьшилось с
524,8 до 140,3 экз. /м2, а биомасса с 2,9 до 2,7 г/м2.
Таблица 14 - Численность и биомасса зообентоса рек
Азово-Кубанской равнины
|
Река
|
Биомасса, г/м2
|
|
Ея
|
389,5
|
2,6
|
|
Челбас
|
456,7
|
3,1
|
|
Кирпили
|
516,3
|
3,4
|
|
Понура
|
324,5
|
2,2
|
|
Албаши
|
298,8
|
1,9
|
|
Ясени
|
372,6
|
2,1
|
|
Бейсуг
|
524,8
|
2,9
|
|
Бейсуг (наши данные)
|
140,3
|
2,7
|
|
Примечание: Сведения по численности и биомассе
зообентоса рек Азово-Кубанской равнины: Ея, Челбас, Бейсуг, Кирпили, Понура,
Албаши, Ясени приведены по работе: Рыбохозяйственное освоение. [2013].
|
При сравнение с другими реками Азово-Кубанской низменности,
река Бейсуг в настоящее время имеет наименьшую численность организмов, что
можно объяснить загрязнением и увеличением антропогенной нагрузки на реку.
Также наблюдается снижение численности зообентоса продольного профиля реки
Бейсуг с прошлыми годами.
Биомасса бентосных организмов реки по сравнению с биомассой
прошлых лет значительно не изменилась. Так же как и в прошлые года она
преобладала над биомассой рек: Ея, Понура, Албаши, Ясени и уступала по биомассе
рекам Челбас и Кирпили.
4.6 Значение
зообентоса в питании массовых видов рыб
Ихтиофауна малых степных рек весьма консервативна и скудна.
Без учета интродуцированных рыб в степных реках в настоящее время отмечено 19
видов: щука, плотва, красноперка, линь, обыкновенный пескарь, уклейка, густера,
лещ, обыкновенная верховка, золотой и серебряный караси, сазан, щиповка,
трехиглая колюшка, бычокцуцик, бычок песочник, окунь, судак, обыкновенный ерш
[Белюченко, 2010].
Наиболее массовыми видами ихтиофауны реки Бейсуг, в спектре
питания которых преобладал зообентос, были серебряный карась, плотва и густера.
Густера (Blicca bjoerkna) - пресноводная рыба
семейства карповых. Длина до 35 см (обычно меньше), масса от 200 г до 1,2 кг за
брюшными плавниками киль, не покрыт чешуёй. Это стайная рыба,
держится у дна среди растительности, часто вместе с лещом. Питается как
растительными, так и животными организмами: молодыми побегами водных растений,
личинками насекомых, моллюсками. Нерест с середины мая до конца июня. Объект
промысла и любительского рыболовства [Атлас пресноводных рыб., 2003].
Обыкновенная плотва (Rutilus rutilus) - стайная рыба, обитающая в
богатых растительностью прибрежных зонах. Рыба достигает небольших размеров
обычно до 25 см (встречаются экземпляры до 45 см). Нерестится в конце
марта - начале апрел. Обыкновенная плотва имеет два подвида (тарань и вобла).
Питается растительной пищей, водными беспозвоночными и детритом [Атлас
пресноводных рыб., 2003].
Серебряный карась - пресноводная оседлая
рыба. Питается планктоном, детритом, водорослями, личинками насекомых, червями
и другими беспозвоночными. Нерест порционный обычно в марте - мае. Серебряный
карась -
объект промыслового и любительского лова [Атлас пресноводных рыб., 2003].
При обловах отбирались наиболее массовые виды рыб реки
Бейсуг, в спектре питания которых преобладал зообентос: серебряный карась,
плотва и густера. Размеры плотвы колебались от 15,0 до 18,1 см, масса от 28,0
до 39,1 г, серебряного карася от 19,0 до 31,2 см и от 125,0 до 490,1 г, густеры
от 14,9 до 19,5 см, массой от 29,4 до 42,2 г.
В таблице 15 приведены данные по соотношению отдельных групп
зообентоса в пищевом спектре массовых видов рыб.
Таблица 15 - Пищевой спектр массовых видов рыб реки Бейсуг в
процентах от массы содержимого кишечника
|
Кормовые объекты
|
Плотва
|
Серебряный карась
|
Густера
|
|
зообентос
|
|
Брюхоногие моллюски
|
0,0
|
0,0
|
27,8
|
|
Разноногие раки
|
24,1
|
8,5
|
0,0
|
|
Жесткокрылые
|
0,0
|
23,7
|
11,1
|
|
Хирономиды
|
29,6
|
30,5
|
50,0
|
|
Стрекозы
|
1,9
|
6,8
|
8,3
|
|
прочие компоненты
|
|
Детрит
|
1,8
|
9,0
|
1,8
|
|
Водоросли
|
7,5
|
1,1
|
1,0
|
|
Копеподы
|
18,5
|
11,9
|
0,0
|
|
Кладоцеры
|
16,6
|
8,5
|
0,0
|
|
Итого
|
100,0
|
100,0
|
100,0
|
Основными компонентами, преобладающими в питания
исследованных нами рыб, являлись личинки хирономид, так же у всех исследованных
рыб встречались стрекозы. Брюхоногие моллюски были выявлены в большом
количестве в питании густеры. Жесткокрылые в наибольшем количестве отмечены в
пищевых комках серебряного карася.
В спектре питания исследованных рыб преобладали бентосные
организмы. Относительная масса зообентоса в кишечниках плотвы составляла 55,6 %,
у серебряного карася 69,5 %, а у густеры 97,2 % от общего количества массы.
Анализ питания рыб исследованной реки Бейсуг показал, что
зообентосные организмы имеют большое значение. Причём, развитие бентических
организмов подвержено значительным сезонным колебаниям, что в свою очередь
отражается на количественном и качественном составе пище бентософагов.
Заключение
На основании проведенных исследований можно сделать следующие
выводы:
. Зообентос реки Бейсуг в летне-осенний период 2014 г.
включал 11 видов зообентосных организмов, относящихся к 12 семействам.
Наибольшим видовым разнообразием характеризовалось нижнее течение реки (10
видов), минимальным - верхнее (четыре вида).
. Численность зообентоса зависела от участка реки. Наиболее
высокие количественные характеристики зообентоса отмечены в верхнем и нижнем
участках водотока - 162,5 и 150,0 экз. /м2 соответственно.
Минимальные количественные показатели в развитии зообентоса выявлены в среднем
течении р. Бейсуг - 108,3 экз. /м2.
. Значение биомассы зообентоса в реке Бейсуг зависело от её
участка. Наиболее высокие значения биомассы всего зообентоса были отмечены в
верхнем и среднем участках водотока - 284,32 и 211,63 г/м2
соответственно, минимальные в нижнем течении р. Бейсуг - 148,35 г/м2.
Биомасса кормового зообентоса имела максимальные в среднем и нижнем участках
реки - 3,30 и 2,52 г/м2, а в верхнем максимальные - 0,98 г/м2.
. На всех участках реки доминирующим по численности
компонентом зообентоса был представитель семейства хирономид - звонец мохнатоусый
(Chironomus plumosus). Его относительная численность изменялась от 19,9
в среднем течении до 49,0 % в нижнем течении реки. Кроме него в нижнем течении
высокой относительной численности достигал бокоплав (Gammarus lacustris)
(19,4 %), в среднем течении катушка роговая (Planorbarius corneus) (19,2
%), в верхнем - перловица (Unio pictorum) (18,0 %).
. Доминирующим компонентом по биомассе на всех участках реки
был представитель двустворчатых моллюсков - перловица. Её относительная
численность изменялась от 99,30 % в нижнем течении до 99,65 % в верхнем течении
реки. Кроме неё в нижнем и среднем течении высокой относительной численности
достигал обыкновенный прудовик (Limnaea stagnalis) - 0,98 и 1,27 %
соответственно.
. Временная динамика количественных показателей зообентоса в
верхнем и нижнем участках реки проявлялась в снижении численности животных от
июля к сентябрю, в среднем - в возрастании. Временная динамика биомассы была
аналогичной.
. В летне-осенний период 2014 г. численность зообентоса в р.
Бейсуг была существенно ниже, чем в других реках Азово-Кубанской равнины, а
показатели биомассы были сопоставимы. Наиболее благополучным состоянием
характеризуется зообентос среднего и нижнего течения р. Бейсуг.
. Зообентос реки Бейсуг играет важную роль в питании массовых
видов рыб этой реки - плотвы, серебряного карася, густеры. Относительная масса
зообентоса в кишечниках изученных рыб изменялась от 55,6 % (плотва) до 97,2 %
(густера).
Библиографический
список
1. Актуальные
вопросы экологии и охраны природы экосистем малых рек: сборник материалов
Межреспубликанской научно-практической конференции. Краснодар, 1992.223 с.
2. Абаев
Ю.И. Современное состояние и перспективы интенсивного рыбохозяйственного
освоения прудов на степных реках Краснодарского края // Актуальные вопросы
экологии и охраны природы экосистем малых рек: сб. науч. тр. Краснодар, 1992.С.
202-204.
. Алимов
А.Ф., Финогенова Н.П. Количественная оценка роли сообществ донных животных в
процессах самоочищения пресноводных водоёмов // Гидробиологические основы
самоочищения вод.Л., 1976. С.146-149.
. Арбанина
И.П., Савицкий Б.П., Рыдный С.А. Бентос мелиоративных каналов Полесья. Минск,
1988.40 с.
. Атлас
пресноводных рыб России: в 2 т. / под ред. Ю.С. Решетникова. М., 2003.
. Баканов
А.И. Мониторинг состояния р. Оки по зообентосу // Экология. 1996. № 22. С.156-160.
. Барышман
Ф.С. Лесоразведение в комплексе мер защиты почв от эрозии. Краснодар, 1983.112
с.
. Безднина
С.Я. Экология мелиорации и водного хозяйства. М., 2001. С.30-31.
. Безматерных
Д.М. Зообентос как индикатор экологического состояния водных экосистем Западной
Сибири. Новосибирск, 2007.87 с.
. Белюченко
И.С. Агроэкология. Краснодар, 1993.132 с.
. Белюченко
И.С. Агроландшафтная экология. Краснодар, 1996.250 с.
. Белюченко
И.С. К вопросу о спецификации речной гидрологии Краснодарского края //
Экологические проблемы Кубани. 2004. №26. С.5-9.
. Белюченко
И.С. Экология Кубани: в 2 ч. Краснодар, 2005. Ч.1.513 с.
. Белюченко
И.С., Мамась Н.Н. Оценка состояния речных систем степной зоны края и
предложения по улучшению их экологической ситуации // Экологические проблемы
Кубани. Краснодар, 2005.С. 198-207.
. Берг
Л.С. Биполярное распространение организмов и ледниковая эпоха: в 5 ч.М., Л.,
1962. Ч.5. С.101-129.
. Болкунов
О.А., Рудакова Н.А., Ерзиков О.О. Краткая характеристика и потенциальная
рыбопродуктивность по зообентосу степных рек // Технологический форсайт: матер.
Всерос. науч. - практ. конф. студентов, аспирантов и молодых учёных. Краснодар,
2014. С.157-160.
. Борисов
В.И. Реки Кубани. Краснодар, 1978.79 с.
. Борисов
В.И. Химическая география речных вод Краснодарского края // Материалы научной
конференции по вопросам географии Кубани. Краснодар, 1971. С.43.
. Борисов
В.И. Реки Кубани. Краснодар, 2005.120 с.
. Вагинов
А.Н. Гидрология рек. Москва, 1976.338 с.
. Вальков
В.Ф. Почвы Краснодарского края, их использование и охрана. Ростов-н/Д.,
1996.192 с.
. Волошина
Г.В. Тяжёлые металлы в пойменных почвах реки Понура // Труды Кубанского
аграрного университета. 2007. № 3.С. 191-195.
. Волошина
Г.В. Микробиологическая оценка реки Понура // Экология и жизнь: матер.9-й
Междунар. науч. - практ. конф. Пенза, 2006.С. 208-210.
. Волошина
Г.В. Экологическая оценка состояния поверхностных вод реки Понура //
Экологический вестник Северного Кавказа. Краснодар, 2006. №1. С.118-122.
. Гайдай
А.А. Некоторые аспекты состояния степных рек Краснодарского края //
Экологические проблемы Кубани. Краснодар, 2005.С. 207-211.
. Гайдай
А.А. Оценка экологического состояния бассейна реки Бейсуг и предложения по
улучшению его функционирования: автореф. дисс. … канд. биол. наук. Краснодар,
2006.180 с.
. Горелов
С.К. Тектонические движения Азово-Кубанской равнины по данным изучения
отложений и морфологии пойм // Материалы Всесоюзного совещания по изучению
четвертичного периода. М., 1961. С.75.
. Данилов
Г.Г. Защитные лесонасаждения и охрана почв. М., 1983.232 с.
. Дегтярёва
В.Н. К вопросу индикации загрязнения природных экосистем хлорорганическими
соединениями // Эколого-токсикологическая характеристика г. Казани и природной
зоны. Казань, 1991. С.38-45.
. Дулепов
В.И. Системная экология. Владивосток, 2004.204 с.
. Жадин
В.И. Методы гидробиологического исследования. М., 1960.72 с.
. Жадин
В.И., Герд С.В. Реки, озера и водохранилища СССР, их фауна и флора, М.,
1961.143 с.
. Жадин
В.И. Жизнь в реках. Бентос // Жизнь пресных вод. М.; Л., 1950. С.149-183.
. Кирейчук
А.Г. Определитель водных жуков Росси: общая часть и определители семейств. М.,
2001.153 с.
. Климатические
изменения зообентоса Азовского моря в условиях антропогенных воздействий // Водные
ресурсы. 2011. №6. С.698-706.
. Комулайнен
С.Ф., Круглова А.Н., Хренников В.В., Широков В.А. Методические рекомендации по
изучению гидробиологического режима малых рек. Петрозаводск, 1989.41 с.
. Коровин
В.И. Экологическая проблема рек Восточного Приазовья и пути ее решения //
Актуальные вопросы экологии и охраны природы Азовского моря и Восточного
Приазовья. Краснодар, 1990. С.218-220.
. Коровин
В.И. Гидрографическая сеть Северного Кавказа. Краснодар, 1981.78 с.
. Кондратьев
Н.Е., Попов И.В., Снищенко Б.Ф. Основы гидроморфологической теории руслового
процесса.Л., 1982.272 с.
. Крылова
А.Г. Водные биоценозы: закономерности формирования и практическое значение.
Краснодар, 1982.100 с.
. Кузьмина
И.А. Малый практикум по гидробиологии. М., 2007.232 с.
. Кутикова
Л.А., Старобогатов Я.И. Определитель пресноводных беспозвоночных европейской
части СССР. М., 1977.130 с.
. Кузнецов
Е.В., Шевченко Г.В., Кадышев С.Л. Подтопление земель и оценка воздействия
степных рек Краснодарского края на прилегающие территории // Актуальные вопросы
мелиорации, гидравлики и экологии. Краснодар, 2000. С.6-10.
. Лакин
Г.Ф. Биометрия: учебное пособие для университетов и педагогических институтов.
М., 1973.343 с.
. Лилиенберг
Д.А. Региональная геоморфология Кавказа. М., 1979.236 с.
. Маккавеев
Н.И., Чалов Р.С. Русловые процессы. М., 1988.264 с.
. Мельникова
Т.Н. Водоносность рек Северо-Западного Кавказа. Майкоп, 2003.132 с.
. Москул
Г.А. Рыбохозяйственное освоение и повышение рыбопродуктивности водоёмов малых
рек Краснодарского края // Рациональное использование и охрана малых рек.
Таллин, 1985. С.36-37.
. Москул
Г.А. Временные рекомендации по рыбохозяйственному освоению и повышению
рыбопродуктивности водоёмов, расположенных на степных реках Краснодарского
края. Краснодар, 1987.14 с.
. Нагалевский
В.Я. Современное состояние прудов (малых водохранилищ) Азово-Кубанской равнины
// Актуальные вопросы экологии и охраны природы экосистем малых рек. Краснодар,
1992. С.137-140.
. Нагалевский
Ю.Я., Щеглова З.П. Экосистемы устьевых областей степных рек Восточного
Приазовья // Актуальные вопросы экологии и охраны природы предгорных экосистем:
матер. межреспубл. науч. - практ. конф. Краснодар, 1993. С.210-215.
. Нежиховский
Р.А. Наводнения на реках Советского союза // Человек и стихия. М., 1989 С.115-119.
. Никаноров
А.Н., Страдомская А.Г. Химический состав органических и минеральных веществ
иловых донных отложений незагрязнённых водных объектов // Водные ресурсы. 2004.
Т.31, №5. С.546-555.
. Одум
Ю. Экология. М., 1986. Т.1.328 с.
. Океанографическая
энциклопедия. М., 1974. С.79-81.
. Панкратова
В.Я. Личинки и куколки комаров подсемейств Podonominae и Tanypodinae фауны СССР
/ Определители по фауне СССР.Л., 1977.154 с.
. Плотников
Г.К. Ихтиофауна различных водных экосистем Северо-Западного Кавказа. Краснодар,
2001.62 с.
. Поляков
Ю.К. К вопросу о донных отложениях оз. Панаярви // Гидрологические исследования
на водоёмах Карелии.Л., 1994.60 с.
. Полоскин
А., Хаитов В. Полевой определитель беспозвоночных. М., 2006.16 с.
. Райнин
В.Е. Экологические проблемы великих рек // Мелиорация и водное хозяйство. 2000.
№2. С.31-34.
. Ресурсы
поверхностных вод СССР. Л., 1973.447 с.
. Руководство
по методам гидробиологического анализа поверхностных вод и донных отложений /
отв. ред.В.А. Абакумова. Л., 1983.240 с.
. Рыбохозяйственное
освоение и способы повышения рыбопродуктивности рек Азово-Кубанской равнины /
Г.А. Москул [и др.] // Рыбное хозяйство: научно-практический и производственный
журнал. 2013. №2. С.79-83.
. Рылов
В.М. Пресноводная фауна. Л., 1930.320 с.
. Свешников
В.А., Алигаджиев М.М. Жизненные формы бентосных беспозвоночных Каспийского
моря. Махачкала, 1989.128 с.
. Свистунов
Ю.А., Кузнецова А.Е., Шевченко Г.В. Проблемы переувлажнения земель в
Краснодарском крае // Актуальные вопросы мелиорации, гидравлики и экологии: сб.
науч. тр. Краснодар, 2000. С.23-30.
. Симов
В.Г. Гидрология устьев рек Азовского моря. М., 1989.326 с.
. Системы
земледелия в Краснодарском крае на 1990-1995 годы и на период до 2000 года.
Краснодар, 1990.146 с.
. Смирнов
Н.Н. Фауна СССР. Ракообразные.Л., 1971.268 с.
. Фильчагов
Л.П. Возрождение малых рек. Киев, 1989.183 с.
. Никаноров
А.М., Страдомская А.Г. Хроническое загрязнение пресноводных объектов по данным
о накоплении пестицидов, нефтепродуктов и других токсичных веществ в донных
отложениях // Водные ресурсы. 2007. №3. С. 337-344.
. Чалов
Р.С., Чернов А.В. Морфология и динамика русел малых рек России и их
антропогенное изменения // Малые реки России. М., 1994. С. 66-80.
. Чередниченко
Л.И. История развития долины реки Кубань // Актуальные вопросы изучения
экосистемы бассейна Кубани. Краснодар, 1988. С. 11-13.
. Чупахин
В.М. Физическая география Северного Кавказа. Ростов - н/Д., 1974.215 с.
. Шарова
И.Х. Зоология беспозвоночных. М., 1999.230 с.
. Яковлев
В.А. Количественная оценка влияния антропогенных факторов на формирование
донных биоценозов субарктического водоёма // Антропогенное воздействие на
экосистемы Кольского Севера. Апатиты, 1988. С.68-79.
. Яшнов
В.А. Практикум по гидробиологии. М., 1969.160 с.