Основы биофизики
Министерство образования Российской
Федерации
Рязанская государственная
радиотехническая академия
Кафедра микроэлектроники
Курс лекций
Основы
биофизики
проф. Вихрова С.П.
по направлению 653900
"Биомедицинская техника"
Лекции составлены по книге "Медицинская биофизика" / Под
редакцией В.О. Самойлова.
Из-во Военно-медицинская академия им. С.М. Кирова. Л. 1986 г.
Общие
сведения и задачи курса
Предметом биофизики является изучение физических и физико-химических
процессов, лежащих в основе жизни. По природе объектов исследования, биофизика
является типичной биологической наукой, а по методам изучения и анализа
результатов исследования является своеобразным разделом физики. Биофизические
методы созданы на основе физических и физико-химических методов изучения
природы. В этих методах должны сочетаться трудно совместимые качества:
. Высокая чувствительность.
. Большая точность.
Этим требованиям не удовлетворяют полностью никакие методы, однако,
наиболее широкое применение получили для биофизических исследований следующие
методы:
оптические;
радиоспектроскопия
ультразвуковая радиоскопия;
электронно-парамагнитная резонансная спектроскопия (ЭПР);
ядерная магнитная резонансная спектроскопия.
Необходимо отметить, что любые исследования требуют, чтобы регистрирующие
приборы не вносили искажений в изучаемый процесс, однако, трудно сравнить
какую-либо физическую систему с живым организмом по необычайно высокой
чувствительности организма к любым воздействиям на него. Воздействия не просто
нарушают нормальный ход биологических процессов, а вызывают сложные
приспособительные реакции, разнообразные в различных органах и в различных
условиях. Искажение смысла измерений может оказаться столь существенным, что
становится невозможно вносить поправки в явления, не свойственные изучаемому
объекту. При этом методы коррекции, используемые с успехом в физике и технике,
зачастую бесполезны в биофизике.
Для лучшего понимания области применения биофизических методов,
рассмотрим основные направления исследований в биофизике:
) молекулярная и квантовая биофизика изучают физическую структуру и
свойства биологически важных молекул, физические процессы, которые обеспечивают
их функционирование, а также исследуют термодинамику биологических систем,
перенос энергии и заряда по биомолекулам, квантово-механические особенности их
строения;
) биофизика клетки связана с физическими и физико-химическими свойствами
клеточных и субклеточных структур, закономерностями их деления, с особенностями
их обмена веществ (метаболизмом), а также, с биофизическими механизмами
специальных функций клеток;
) биофизика органов чувств скрывает физические и физико-химические
механизмы восприятия раздражителя рецепторами аппарата сенсорных систем
человека и животных на квантовом, молекулярном или клеточном уровнях;
) биофизика сложных систем изучает проблемы межклеточного взаимодействия,
передачи информации в биологических каналах связи и управления функциями живых
организмов;
) биофизика внешних воздействий изучает механизмы воздействия на организм
физических факторов среды (например, полей).
1. Биоэнергетика. Первое начало термодиамики (общие сведения)
Жизненные процессы, при всем многообразии, имеют и общие черты, в
частности, любой из процессов требует затрат энергии. В этой связи важным
направлением биофизических исследований является изучение преобразования
энергии в биологических системах. Процессы энергообеспечения организма за счет
внешних энергетических ресурсов, составляют предмет исследования биоэнергетики.
В биоэнергетике выделены два подхода:
) исследуются механизмы энергетических процессов, протекающие в организме
на молекулярном и субмолекулярном уровнях;
) изучаются особенности биологических процессов на основе общих законов
превращения энергии, без детального изучения их молекулярных механизмов. Это
составляет содержание биологической термодинамики.
В термодинамике объектом исследования служит система, под которой
понимают совокупность объектов, ограниченных в той или иной степени от
окружающей среды. Различают изолированные системы, которые не обмениваются
энергией, веществом и информацией с окружающей средой и открытые системы, где
такой обмен происходит. Живой организм относится к открытой системе.
Состояние любой системы характеризуется некоторыми параметрами. Одни из
них не зависят от массы или числа частиц в системе, то есть, от размеров,
другие параметры пропорциональны этим аргументам. Первые получили название
интенсивных термодинамических параметров, к ним относятся: температура,
давление и т.д. Параметры второй группы называются экстенсивными
термодинамическими параметрами. Например, это объем, энергия, энтропия и т.д.
Энергию системы можно представить, состоящей из двух частей:
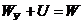
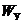 - энергия системы, как целого;
- энергия системы, как целого;
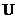 - внутренняя энергии (энергия атома и т. д.).
- внутренняя энергии (энергия атома и т. д.).
Смысл первого начала термодинамики сводится к тому, что изменение внутренней
энергии системы может произойти только при обмене энергией с окружающей средой.
Энергетический обмен между системой и средой осуществляется двумя способами:
посредством передачи тепла и (или) совершением работы.
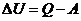
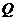 - количество тепла;
- количество тепла;
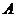 - работа.
- работа.
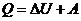 - первое начало термодинамики.
- первое начало термодинамики.
Знак в формуле принимает следующие образы:
положительным считают то тепло, которое получает система из окружающей
среды;
работу считают положительной, когда система производит ее над окружающими
телами.
Рассмотрим некоторые способы совершения работы:
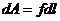 - механическая работа.
- механическая работа.
 - работа при постоянном объеме.
- работа при постоянном объеме.
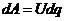 - работа при перемещении заряда и разность потенциалов.
- работа при перемещении заряда и разность потенциалов.
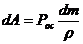 -
- 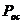 - осмотическое давление.
- осмотическое давление.
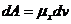 - изменение числа молей,
- изменение числа молей,
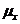 - химический потенциал.
- химический потенциал.
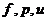 ... - величины, вызывающие причины действия работы,
интенсивные параметры.
... - величины, вызывающие причины действия работы,
интенсивные параметры.
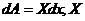 - обобщающая сила, которая вызывает работу;
- обобщающая сила, которая вызывает работу;
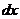 - обобщающая координата.
- обобщающая координата.
 , ... - экстенсивные параметры.
, ... - экстенсивные параметры.
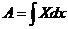 ,
, 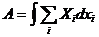
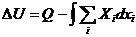
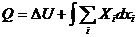
Количество тепла, получаемое системой, определяется изменением внутренней
энергии системы, а также суммой всех видов работы, совершенной системой.
2. Свободная
и связанная эенергия
Движение частиц в любом теле может быть упорядоченным и неупорядоченным.
Например, у всех молекул газа (или воды), когда он течет по трубе, есть общая
составляющая скорости, которая определяет движение газа, как целого. Такое
движение называют упорядоченным. Кроме того, молекулы газа участвуют в
непрерывном неупорядоченном тепловом движении. Упорядоченное движение - электрический
ток, а неупорядоченное движение - тепловое движение электронов газа.
Между упорядоченным и неупорядоченным движениями существует
принципиальная разница: упорядоченное движение может полностью превратиться в
неупорядоченное, а переход из неупорядоченного движения в упорядоченное никогда
не бывает полным.
Причина различия между двумя такими движениями связана с неодинаковой
вероятностью каждого из них.
Для обеспечения упорядоченного движения необходимо, чтобы все частицы
имели в данный момент одинаковые по величине и направлению составляющие
скорости, а когерентные волны должны иметь одинаковую частоту и фазу.
Такое состояние является, менее вероятно, чем-то, при котором скорости
частицы или фазы волн различны.
Внутренняя энергия в идеальном газе полностью, а в других телах частично,
связана с неупорядоченным тепловым движением молекул. В то же время, совершение
работы всегда требует переноса вещества, то есть, упорядоченного движения.
Поэтому принципиально невозможно всю внутреннюю энергию тела использовать
для совершения работы.
Только та часть внутренней энергии системы, которую в принципе можно
использовать для совершения работы, называют свободной энергией - G. Остальную
часть внутренней энергии нельзя превратить в работу, и ее называют связанной
энергией.
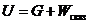
Следовательно, работа , совершенная системой в любом процессе, не может быть
больше, чем изменение 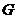 этой системы.
этой системы.
3. Обратимые и необратимые процессы
Те процессы, в которых 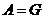 , называются обратимыми, так как, пустив такой процесс в
обратном направлении и затратив работу, можно вернуть систему в исходное
состояние.
, называются обратимыми, так как, пустив такой процесс в
обратном направлении и затратив работу, можно вернуть систему в исходное
состояние.
Таких процессов в природе не существует. Все реальные процессы -
необратимы.
Иными словами, не может быть полностью преобразовано в .
При таком преобразовании часть обязательно превращается в тепло. Для
достижения максимальной степени обратимости термодинамических процессов нужно
добиваться минимальной разности между и.
И в технике, и в биологии представляет интерес в первую очередь , совершенная системой, поэтому важно
знать не столько полную 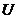 системы, сколько ее .
системы, сколько ее .
системы, как является функцией в которой находится система. Наиболее
важными параметрами являются:
температура;
давление;
число молей вещества;
а при наличии электрического поля, и его напряженность 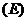 .
.
Тогда,
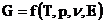
Во многих системах, в частности, в живых организмах, наиболее важным
источником 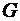 является химическая энергия молекул, входящих в систему.
является химическая энергия молекул, входящих в систему.
В этой связи введено понятие химического потенциала.
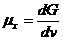
Химический потенциал системы по отношению к конкретному веществу равен
приросту системы при увеличении количества
этого вещества на один моль.
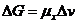 ,
, 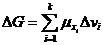
4. Применение первого начала термодинамики к живым организмам
В отличие от тепловых машин, живые организмы производят не за счет тепловой энергии, а за
счет использования химической энергии пищевых продуктов, усвоенных ими. В этой
связи уравнение, согласно которому изменение системы равно ее обмену энергии с
окружающей средой, имеет вид:
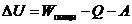 ,
,
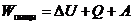
Организм животных имеет постоянную температуру, и химический состав его в
среднем не изменяется, поэтому такого организма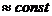 . Следовательно, изменение
. Следовательно, изменение 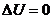 . Тогда данное уравнение имеет вид:
. Тогда данное уравнение имеет вид:
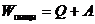
Поскольку существует множество видов работ и обмена тепла с окружающей
средой, то уравнение можно представить:
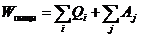
первое начало термодинамики применительно к живым организмам.
Следует заметить, что первичным источником энергии служит Солнце.
Мощность солнечного излучения примерно составляет 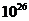 Вт, но только небольшая ее часть,
примерно
Вт, но только небольшая ее часть,
примерно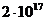 Вт достигает поверхности Земли, а из
этой части, 0,02% поглощается зелеными растениями и запасается ими в процессе
фотосинтеза. Следовательно, поток энергии, извлекаемый зелеными растениями из
солнечного света, имеет порядка
Вт достигает поверхности Земли, а из
этой части, 0,02% поглощается зелеными растениями и запасается ими в процессе
фотосинтеза. Следовательно, поток энергии, извлекаемый зелеными растениями из
солнечного света, имеет порядка 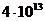 Вт. За счет этой энергии работают все тепловые машины и
осуществляются все процессы жизнедеятельности.
Вт. За счет этой энергии работают все тепловые машины и
осуществляются все процессы жизнедеятельности.
Однако, способы преобразования в работу солнечной энергии,
аккумулированной зелеными растениями в форме химической энергии, в принципе не
одинаковы в тепловых машинах и биологических системах. Различия
термодинамических процессов можно рассмотреть следующей схемой:
В тепловой машине:
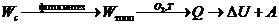
В биологической системе:

Как уже отмечалось, источником для всех живых существ служит
Солнце. Земные растения (аутотрофы) за счет фотосинтеза, создают в течение года
примерно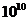 тонн питательных веществ.
Гетеротрофы сами не могут питаться светом, они получают , поедая друг друга или питаясь
растениями. Пищеварение обеспечивается поступлением в клетки продуктов
гидролиза пищи, то есть, углеводов, белков, жиров, в которых заключена солнечного света.
тонн питательных веществ.
Гетеротрофы сами не могут питаться светом, они получают , поедая друг друга или питаясь
растениями. Пищеварение обеспечивается поступлением в клетки продуктов
гидролиза пищи, то есть, углеводов, белков, жиров, в которых заключена солнечного света.
Основным способом использования питательных веществ организмом
является их биологическое окисление. Оно происходит главным образом на
внутренней мембране митохондрий, где сосредоточены ферменты, катализирующие
биологическое окисление (клеточное дыхание). Поэтому митохондрии часто называют
энергетическим цехом клетки.
Энергия, извлекаемая из химических связей питательных веществ при их
биологическом окислении, в некоторых случаях может быть непосредственно
использована для осуществления жизнедеятельности, но основная ее часть идет на
синтез так называемых макроэнергетических соединений, среди которых наиболее
важным является АТФ.
Энергия, запасенная в макроэргах, используется организмом для совершения
различных видов работ, причем механическая (мышечная работа) не является самой
энергоемкой. В жизни человека огромные затраты энергии идут на синтез сложных
биомолекул. Так, для синтеза одного моля белка, требуется от 12000 до 200 тыс.
кДж . "В сборке" одной молекулы
белка участвуют от 1000 до 16 тыс. молекул АТФ (КПД примерно 40 %) Для синтеза
молекулы РНК необходимо примерно 6 тыс. молекул АТФ, еще большей энергии
требуется для образования ДНК, так, на создание одной молекулы ДНК тратится 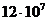 молекул АТФ. Однако, количество
синтезирующихся молекул белка значительно больше, чем нуклеиновых кислот, в
силу разнообразия его функций и постоянного быстрого обновления. В результате,
именно синтез белка в организме наиболее энергоемок, по сравнению с другими
биосинтетическими процессами, так, в течение каждого часа жизни у млекопитающих
белок клеток обновляется примерно на 1%. А белки-ферменты на 10%. У человека,
массой 70 кг, ежечасно обновляется примерно 100 грамм белка.
молекул АТФ. Однако, количество
синтезирующихся молекул белка значительно больше, чем нуклеиновых кислот, в
силу разнообразия его функций и постоянного быстрого обновления. В результате,
именно синтез белка в организме наиболее энергоемок, по сравнению с другими
биосинтетическими процессами, так, в течение каждого часа жизни у млекопитающих
белок клеток обновляется примерно на 1%. А белки-ферменты на 10%. У человека,
массой 70 кг, ежечасно обновляется примерно 100 грамм белка.
Другой важной "статьей" расхода в организме является поддержание
физико-химических градиентов на клеточных мембранах. Внутри живой клетки
концентрация ионов и вещества, отличается от межклеточной среды, то есть, на
клеточной мембране существует градиент концентрации. Различие концентрации
ионов и молекул вызываются появлением и других градиентов: остматического,
электрического, фильтрационного и т.д.
Наличие градиентов вызывает непрерывный перенос вещества через клетки
мембран (пассивный транспорт). Пассивный транспорт должен был бы уменьшить
величину градиентов, то есть, выровнять концентрацию и другие физико-химические
параметры. Однако, в нормальных условиях функционирования клетки, градиенты на
мембране стабильно поддерживаются на определенном уровне, что обусловлено
способностью биологической системы переносить вещества против градиентов. Такой
транспорт называется активным транспортом. Активный транспорт нуждается в
затратах на него, которое в большинстве
случаев черпается из АТФ следовательно активный транспорт представляет собой
одну из форм работы биологической системы с КПД примерно 20-25%. КПД мышечного
сокращения организма не превышает 20%.
. Тепловой баланс организма. Способы теплообмена
Наряду с совершением работы , организм преобразует питательного вещества в . В конечном итоге, вся энергия,
полученная организмом с пищей, кроме той части, которая идет на механическую
работу, превращается в тепловую, и в виде тепла отдается организмом в окружающую
среду. Принято выделять несколько этапов в этом теплообразовании, которые
делятся на два вида:
. Прежде всего, тепловые потери присущи биологическому окислению
питательных веществ, в ходе которых синтезируется АТФ. Выделяющаяся при этом
тепловая энергия называется первичным теплом.
. Все остальные теплообразования называют вторичным теплом. К ним
относят: тепло при синтезе макромолекул (кроме АТФ); при поддержании градиентов
за счет активного транспорта; при мышечном сокращении; при трении мышц, кровеносных
сосудов, суставов и т. д.; при распаде белков и других макромолекул; при
пассивном транспорте веществ.
На рисунке показана схема преобразования солнечной энергии в организме
человека.
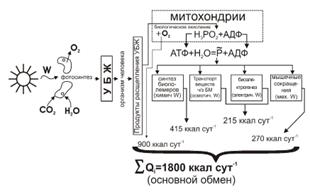
Вся тепловая энергия, образующаяся в организме человека, уходит из него.
Теплообмен осуществляется на поверхности тела, при этом различают четыре
основных его способа:
. Теплопроводность - 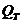
. Конвекция - 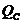
. Излучение - 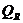
. Испарение - 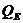
1. Теплопроводность играет основную роль
в переносе тепла через одежду. Тепловая энергия, переносимая посредством
теплопроводности, может быть вычислена по следующей формуле:
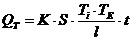
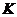 - коэффициент теплопроводности;
- коэффициент теплопроводности;
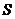 - площадь теплообмена;
- площадь теплообмена;
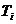 - температура поверхности тела;
- температура поверхности тела;
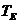 - температура окружающей среды;
- температура окружающей среды;
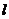 - толщина слоя (одежды);
- толщина слоя (одежды);
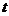 - время, в течение которого идет процесс теплообмена.
- время, в течение которого идет процесс теплообмена.
. Конвекцией называют перенос тепла, перемещающийся средой, то есть,
движущийся газом или жидкостью. Различают естественную и принудительную
конвекцию. При естественной конвекции причиной перемещения среды является сама
разность температур в различных ее частях. Например, холодный воздух, как более
плотный и тяжелый, опускается вниз и вытесняет легкий теплый воздух. В случае
принудительной конвекции (ветер, вентилятор) имеется внешняя сила,
принудительная конвекция значительно эффективнее переносит тепло, чем
естественная. Перенос тепла при конвекции описывается такой же формулой, как и
для теплопроводности, но уже имеет не постоянное значение, а зависит от конкретных
условий, в которых находится организм.
3. Перенос тепла излучением осуществляется путем испускания инфракрасных
лучей. В соответствии с законом Вина, максимальное излучение при температуре
поверхности человеческого тела 36,7 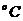 , приходится на длину волны 10
микрон. Величину энергии, излучаемую телом, можно приблизительно найти по
формуле, полученной из закона Стефана-Больцмана:
, приходится на длину волны 10
микрон. Величину энергии, излучаемую телом, можно приблизительно найти по
формуле, полученной из закона Стефана-Больцмана:
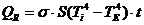
 - так как, для данной инфракрасной (ИК) области спектра,
человек представляет собой абсолютно черное тело.
- так как, для данной инфракрасной (ИК) области спектра,
человек представляет собой абсолютно черное тело.
. Тепло, отводимое от организма путем испарения, рассчитывается по
формуле:
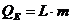
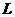 - скрытая теплота (удельная) испарения
- скрытая теплота (удельная) испарения 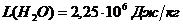
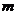 - масса жидкости, испаряемая с поверхности тела.
- масса жидкости, испаряемая с поверхности тела.
Теплоотдача осуществляется только при испарении жидкости с поверхности
тела. Если человек выделяет много пота, но условий для его испарения нет, то
теплоотдача не является эффективной. Так, при 100 % относительной влажности
(паровая баня), испарение полностью прекращается. С кожных покровов человека
испаряется не только вода, но и межклеточная жидкость. За сутки с поверхности
кожи испаряется примерно 0,3-0,4 л жидкости. Жидкость испаряется не только с
кожи, но и со слизистых оболочек. Так, у человека в сутки испаряется 0,3-0,4 л
жидкости со слизистых оболочек дыхательных путей.
Испарение является наиболее эффективным способом теплообмена организма
при высокой температуре и низкой влажности внешней среды. Все остальные способы
теплопередачи функционируют только тогда, когда температура окружающей среды
ниже, чем температура кожи человека, в противном случае они превращаются в
механизм дополнительного нагрева организма. Это обстоятельство учитывается
уравнением теплового баланса организма человека, которое имеет вид:
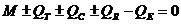
М - теплопродукция (количество тепла, которое выделяется в организме в
сутки). В уравнении знак "+"- если температура окружающей среды
больше то есть 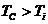 , знак "-" - если
, знак "-" - если 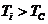 .
.
. Химическая и физическая терморегуляция
Процессы теплообмена очень важны для жизнеобеспечения организма.
Поддержание постоянства температуры организма (температурный гомеостазис)
является необходимым условием жизни человека.
В этой связи все процессы, отображенные в уравнении теплового баланса,
имеют надежную регуляцию. Различны механизмы химической и физической
терморегуляции.
Под химической терморегуляцией понимают возможные усиления или ослабления
теплопродукции (М) за счет изменения интенсивности окислительных процессов.
Химическая терморегуляция обеспечивается влиянием на обмен веществ
(метаболизм). Сам по себе метаболизм очень важен для организма, и его изменение
в условиях поддерживаемой определенной температуры, является крайне
нецелесообразным. Организм прибегает к химической терморегуляции только в
особых случаях.
В обычных условиях основным способом поддержания температуры является
физическая терморегуляция, то есть, регуляция температуры за счет следующих
механизмов теплоотдачи: 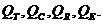
Теплообмен организма с окружающей средой происходит на поверхности тела.
Коэффициент теплопроводности живых тканей имеет низкое значение, поэтому роль
теплопроводности в отведении тепловой энергии от внутренних органов к поверхности
кожи и к слизистым оболочкам - невелика.
Основное значение в этом процессе, то есть, в обеспечении терморегуляции
внутренних органов, принадлежит кровообращению. Теплоемкость крови достаточно
большая (как у воды), и нормальный кроваток достаточен для эффективного отвода
тепла от внутренних органов к поверхности тела. Регуляция такого теплопереноса
осуществляется главным образом за счет усиления или ослабления кровотока.
Другими словами, посредством сосудистых реакций. При необходимости отдать
большее количество тепла, кровеносные сосуды кожи и слизистых оболочек
расширяются, что приводит к значительному увеличению массы циркулирующей в них
крови, имеющей температуру внутренних органов, при этом возрастает и
теплоотдача. Для уменьшения теплоотдачи происходит сужение кровеносных сосудов.
У человека значительные потери тепла происходят через кисти рук и стопы
ног. Так, при переходе от холода к теплу, кровообращение в руке человека
увеличивается в 30 раз, а в пальцах - примерно в 600 раз.
Испарение является наиболее эффективным способом теплообмена организма
при высокой температуре. Физическое терморегулирование является многофакторной
системой, которая весьма эффективно обеспечивает постоянство температуры
организма. Многофакторность позволяет регулировать температуру тела в различных
условиях, при исключении одних механизмов, работают другие.
В биофизике, физиологии и медицине теплопродукцию принято называть
энерготратами организма. Энерготраты сильно изменяются в зависимости от
условий, в которых находится организм, и в зависимости от характера его
деятельности, так как все это влияет на обмен веществ. Для оценки
функционального состояния организма необходимо создание стандартных условий при
измерении его энерготрат, то есть, при измерении величины тепловой энергии,
выделяемой организмом в окружающую среду. За стандартные условия приняты такие,
при которых энерготраты организма минимальны. Для этого нужно исключить влияние
тех факторов, которые усиливают энергообмен, в частности: мышечная работа,
прием пищи, эмоциональное напряжение, отклонение температуры и влажности за
пределы зоны комфорта и т.д.
Величину основного обмена (энерготрат) целесообразно измерять в состоянии
бодрствования (не во время сна), но пациент должен спокойно лежать в постели.
Процедуру рекомендуется проводить рано утром (5-6 часов утра), когда, в
соответствии с суточным ритмом, интенсивность метаболизма самая низкая. За двое
суток до измерения из рациона пациента исключается животная белковая пища.
Измерение проводят натощак, то есть, через 12-14 часов после последнего приема
пищи, при этом температура в помещении должна быть в пределах 20-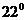 С, а относительная влажность -
50-60%. Величина теплопродукции измеряется несколько раз для получения
статистически достоверного результата.
С, а относительная влажность -
50-60%. Величина теплопродукции измеряется несколько раз для получения
статистически достоверного результата.
Так, у здорового мужчины среднего возраста (40-50 лет), средней массой 70
кг, основной обмен составляет 7800 кДж или 1800 ккал за сутки, или 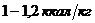 ч, что соответствует мощности - 90
Вт. Величина обмена с единицы поверхности человеческого тела составляет
примерно
ч, что соответствует мощности - 90
Вт. Величина обмена с единицы поверхности человеческого тела составляет
примерно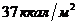 ч, или 100
ч, или 100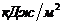 ч. У женщин основной обмен на 7-10%
меньше. Основной обмен сильно зависит от возраста. У новорожденного он
составляет 300ч, а к 70-80 годам понижается до 120ч.
ч. У женщин основной обмен на 7-10%
меньше. Основной обмен сильно зависит от возраста. У новорожденного он
составляет 300ч, а к 70-80 годам понижается до 120ч.
Для определения энерготрат необходимо измерить количество тепла,
выделяемое организмом в окружающую среду за определенный промежуток времени.
Для этого применяют два метода: прямую и непрямую калориметрию.
В методе прямой калориметрии используются специальные физиологические
калориметры, сконструированные таким образом, что в них можно помещать на
нужное время животных или человека. Однако далеко не всегда можно реализовать
прямую калориметрию. В частности, при изучении энергозатрат в ходе трудовой
деятельности.
Чаще всего используют непрямую калориметрию. Этот метод основан на
исследовании газообмена организма. Установлено, что между объемом потребляемого
организмом 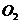 и энерготратами существует линейная зависимость при
фиксированных условиях. Коэффициентом служит так называемый калориметрический
эквивалент , равный количеству тепла, которое образуется в организме при
использовании 1 л для окисления питательных веществ.
и энерготратами существует линейная зависимость при
фиксированных условиях. Коэффициентом служит так называемый калориметрический
эквивалент , равный количеству тепла, которое образуется в организме при
использовании 1 л для окисления питательных веществ.
Калориметрический эквивалент неодинаков при окислении Ж, Б, У, а то,
какие вещества преимущественно окисляются в каждом конкретном случае, можно
определить по дыхательному коэффициенту, который определяется, как объем
выделяемого углекислого газа к объему поглощенного кислорода за один и тот же
промежуток времени.

Так, при преимущественном окислении углеводов дыхательный коэффициент
(ДК) стремится к 1, а при окислении жиров, он имеет самые низкие значения,
примерно 0,7. Существуют специальные таблицы и монограммы, при помощи которых
можно определить величину кислородного эквивалента по найденному значению
дыхательного коэффициента.
Таким образом, посредством газоанализа измеряются объемы поглощенного
кислорода и выделяемого углекислого газа за одно и то же время. Взяв отношение
второго к первому рассчитывают ДК. По ДК находят кислородный эквивалент.
Умножив его на объем поглощаемого кислорода, определяют энерготраты за время
эксперимента.
7. Второе начало термодинамики. Понятие энтропии
Тепловая энергия, образующаяся в организме, представляет собой конкретную
форму, связанной энергии 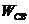 биологической системы. В условиях жизнедеятельности не может быть преобразовано ни в одну
из форм работ, которые совершаются организмом. зависит от степени неупорядоченного
движения молекул или атомов, при этом, ее количественной мерой является
температура. Связь между и температурой - пропорциональна. Однако, величинав различных системах, имеющих
одинаковую температуру, не всегда одинакова. Следовательно, степень неупорядоченного
молекулярного движения зависит не только от температуры, но и еще от каких-то
свойств системы. Эти свойства Клаузис в 1865 г. выразил в виде коэффициента,
который обозначил S и назвал "энтропией". Тогда
биологической системы. В условиях жизнедеятельности не может быть преобразовано ни в одну
из форм работ, которые совершаются организмом. зависит от степени неупорядоченного
движения молекул или атомов, при этом, ее количественной мерой является
температура. Связь между и температурой - пропорциональна. Однако, величинав различных системах, имеющих
одинаковую температуру, не всегда одинакова. Следовательно, степень неупорядоченного
молекулярного движения зависит не только от температуры, но и еще от каких-то
свойств системы. Эти свойства Клаузис в 1865 г. выразил в виде коэффициента,
который обозначил S и назвал "энтропией". Тогда 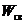 :
:
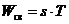 откуда
откуда 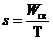 ,
, 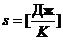
Энтропия - это физическая величина, характеризующая значение данной системы, приходящаяся на
единицу температуры. Тогда, уравнение для первого начала термодинамики
приобретает вид:
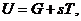
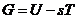 -
- 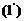 .
.
Более детальный анализ первого начала термодинамики показывает, что
уравнение (1) справедливо только для случая, когда температура системы
постоянная, и объем системы - постоянен. Если объем системы меняется, то первое
начало термодинамики переписывается так:
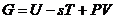 (T- const, V - меняется) -
(T- const, V - меняется) - 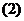 ,
,
При T=const и V=const уравнение (1) имеет вид:
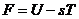
В организме человека температура постоянна, а изменения 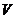 , как правило, незначительны. В этой
связи, при рассмотрении большого жизненного процесса, можно использовать
понятие по Гельмгольцу (уравнение (
, как правило, незначительны. В этой
связи, при рассмотрении большого жизненного процесса, можно использовать
понятие по Гельмгольцу (уравнение (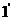 )). Однако, при рассмотрении
процессов, связанных с газообменом, необходимо использовать понятие по Гиббсу (уравнение (2)). На
практике представляет интерес не сама по себе , а ее изменения. Тогда уравнение () и (2) можно записать:
)). Однако, при рассмотрении
процессов, связанных с газообменом, необходимо использовать понятие по Гиббсу (уравнение (2)). На
практике представляет интерес не сама по себе , а ее изменения. Тогда уравнение () и (2) можно записать:
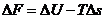
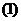
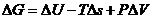
Состояние любой системы характеризуется макроскопическими параметрами,
такими, как температура, давление, объем и т.д. Однако каждому состоянию
соответствует большое число возможных значений параметров, характеризующих
положение и скорости молекул, входящих в данную систему. В термодинамике их
называют микроскопическими параметрами. Если поменять местами две идентичные
молекулы, входящие в состав данной системы, то макроскопические параметры
системы в целом не изменятся. Вероятность состояния системы, определяется
степенью ее упорядоченности. Состояния, для которых характерна высокая
упорядоченность, имеют относительно низкую вероятность. Мало упорядоченные
состояния имеют высокую вероятность существования. С другой стороны, степень
упорядоченности системы характеризуется ее энтропией. Следовательно, между
энтропией состояния и его вероятностью существует зависимость, которую Больцман
выразил формулой:
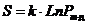
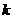 - постоянная Больцмана
- постоянная Больцмана 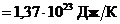
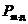 - термодинамическая вероятность, то есть, число возможных
микроскопических состояний, которым может быть реализовано данное
макроскопическое состояние.
- термодинамическая вероятность, то есть, число возможных
микроскопических состояний, которым может быть реализовано данное
макроскопическое состояние.
В реальных телах число молекул огромно, следовательно, количество
вариантов каждого состояния очень велико, и термодинамическая вероятность
выражается огромными числами 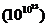 .
.
точно подсчитать можно только в простых случаях. Согласно
формуле Больцмана, при повышении возрастает энтропия. Вместе с тем,
высокой вероятности характерна большая энтропия, к которой стремится любая
система, если на нее не действуют внешние силы. Таким образом, энтропия
указывает направление естественного процесса. В изолированной системе эти
процессы приводят к возрастанию энтропии. При этом доля, связанной энергии системы
возрастает, а доля свободной энергии системы - уменьшается.
Изменение энтропии системы можно выразить через обобщение силы и
обобщение координаты, которые характеризуют данную систему. Уравнение имеет
вид:
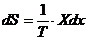
Отсюда легко найти скорость изменения энтропии. Эту величину называют
диссипативной функцией, то есть:
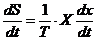
Если в системе действует несколько процессов, то диссипативная функция
имеет вид:

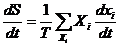
Диссипативную функцию рассчитывают на единицу объема, называют удельной
продукцией энтропии - 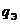 :
:
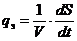
Эту функцию считают количественной мерой необратимости процесса, то есть,
чем больше , тем дальше процесс находится от обратимого.
. Упорядоченость структур в свете второго начала термодинамики
Преобладающей тенденцией эволюции материи на всех уровнях ее
существования, от скопления галактик до атомов, является стремление к
организации и образованию структур. Причина этого заключается в том, что в
любой реальной системе заключена сила взаимодействия ее частиц и, кроме того,
система обычно подвергается действию внешних сил. При наличии таких сил, более
выгодной в энергетическом отношении, как правило, является не хаотическая, а
упорядоченная структура, так как она обеспечивает системе минимум свободной
энергии. Так, при кристаллизации, энтропия системы уменьшается, но одновременно,
значительно снижается внутренняя энергия, так как все ионы и атомы попадают в
положение соответствующее минимальному значению энергии взаимодействия. Однако,
сказанное справедливо при сравнительно низких температурах. Когда в выражение
свободной энергии первый член вносит более существенный вклад, чем второй -
энтропийный. При повышении температуры значение энтропийного фактора
возрастает, а при определенной температуре, его роль становится преобладающей.
Вот тогда термодинамически выгодно оказывается неупорядоченное состояние. Что
проявляется в плавлении кристалла, то есть, изменяется агрегатное состояние
вещества.
Расчет показывает, что у вируса табачной мозаики (ВТМ) будет минимальным при
спиральной структуре. Именно такое строение обеспечивает ВТМ минимально
свободную энергию, что и наблюдается в реальных условиях. Для других вирусов
оказываются более выгодны другие формы структур (сферические, цилиндрические и
т. д.)
Сочетание генетических и термодинамических факторов в процессе роста и
развития характерно для всех живых организмов, вплоть до высших.
Полное изменение энтропии открытой системы можно представить в виде двух
частей:
причиной первой из них служат внутренние процессы, которые необратимы и
сопровождаются выделением энергии;
вторая часть обусловлена обменом энергии и веществом между системой и
окружающей средой.
Тогда изменение энтропии можно представить:
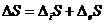
Аналогично можно записать изменение полной свободной энергии для открытой
системы:
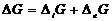
Так как все реальные процессы в открытой системе необратимы, то 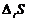 всегда больше 0, а
всегда больше 0, а 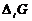 всегда меньше 0, что касается
всегда меньше 0, что касается 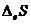 и
и 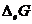 , то знаки этих величин в различных
ситуациях могут иметь разные значения. В ходе обмена с окружающей средой
свободная энергия системы () может и увеличиваться и уменьшаться. В организме первый
случай имеет место при усвоении пищи, а второй - при неблагоприятных
воздействиях на организм, которые вызывают дополнительные затраты . Необходимо отметить, что накопление
в открытой системе (и,
соответственно, уменьшение ее энтропии) всегда сопряжено с возрастанием
энтропии в окружающей среде, то есть, в других телах, с которыми связана данная
система.
, то знаки этих величин в различных
ситуациях могут иметь разные значения. В ходе обмена с окружающей средой
свободная энергия системы () может и увеличиваться и уменьшаться. В организме первый
случай имеет место при усвоении пищи, а второй - при неблагоприятных
воздействиях на организм, которые вызывают дополнительные затраты . Необходимо отметить, что накопление
в открытой системе (и,
соответственно, уменьшение ее энтропии) всегда сопряжено с возрастанием
энтропии в окружающей среде, то есть, в других телах, с которыми связана данная
система.
В термодинамических открытых системах необходимо учитывать непрерывное
изменение и энтропии, обусловленные связью такой системы с постоянно
меняющейся внешней средой, поэтому, при формулировании второго начала
термодинамики для открытых систем, целесообразно ввести величину скорости
изменения энтропии, которая будет определяться следующим выражением:
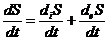 ,
,
где
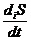 - называется продукцией энтропии;
- называется продукцией энтропии;
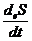 - называется потоком энтропии.
- называется потоком энтропии.
На основе данного выражения, можно сформулировать второе начало
термодинамики для открытых систем:
в открытых системах внутреннее изменение энтропии всегда положительно, а
внутреннее изменение всегда отрицательно.
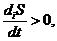
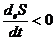 .
.
Этим подчеркивается необратимость реальных термодинамических процессов в
закрытых системах.
Для изолированной системы:
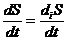 ,
, 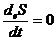
Для поддержания жизни необходимо непрерывное поступление в организм из окружающей среды, чтобы пополнить
постоянную убыль самого организма, идущей на выполнение работы во всех ее
видах и поддержания энтропии организма постоянной. Потребление пищи
обеспечивает с точки зрения термодинамики постоянный приток в организм . Так, за 60 лет жизни человек
съедает примерно 14 тонн углеводов, примерно по 2,5 тонны белков и жиров, и выпивает
примерно 56 тонн воды.
9.
Стационарное состояние
Стационарным называют такое состояние открытой системы, при котором
основные макроскопические параметры системы остаются постоянными. Необходимы
различные стационарные состояния от равновесного состояния.
10. Отличительные признаки стационарного и равносеного состояния
|
Равновесное
|
Стационарное
|
|
1. и работоспособность системы минимальные.
|
1. и работоспособность системы постоянны, но не
минимальны.
|
|
2. Энтропия в системе
максимальна.
|
1. Энтропия в системе
постоянна, за счет равенства продукции и потока энтропии. 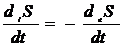
|
|
3. Отсутствие градиентов в
системе.
|
3. Наличие постоянных
градиентов в системе.
|
В состоянии равновесия в системе прекращаются все процессы, кроме
теплового движения молекул, при этом выравниваются все градиенты.
В стационарном состоянии идут химические реакции, диффузия, перенос ионов
и другие процессы, но они так стабилизированы, что состояние системы в целом не
изменяется. В стационарном состоянии существуют градиенты между отдельными
частями системы, но они сохраняют постоянные значения. Это возможно только при
условии, что система из окружающей среды получает вещество и , а отдает продукты реакции и
выделяющееся тепло. Термодинамическим условием стационарного состояния является
равенство между продукцией энтропии, произведенной организмом, и потоком
энтропии, то есть:
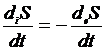 ,
,
тогда полное изменение энтропии равно 0 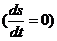 .
.
Термодинамика открытых систем позволяет вскрыть еще одну причину
целесообразности стационарного состояния для биологической системы, которая
сформулирована в теореме Пригожина:
"В стационарном состоянии продукция энтропии имеет постоянное и
минимальное из всех возможных значений, то есть":

Теорема Пригожина показывает, что в стационарном состоянии диссипация происходит с меньшей скоростью, чем
в любых других состояниях, стало быть, в стационарном состоянии системы расходуется наиболее
экономно и поэтому требуется минимальная компенсация ее затрат, то есть, КПД системы
в стационарном состоянии максимален.
Необходимо отметить, что теорема Пригожина справедлива только для таких
состояний, которые мало отличаются от стационарных. В этом случае, скорости
всех процессов выражаются линейными уравнениями (уравнения 1 порядка). Теорема
Пригожина дает термодинамические критерии эволюции линейным системам, которые
формулируются следующим образом:
"Открытая линейная система, если она не находится в стационарном
состоянии, будет изменяться до тех пор, пока скорость продукции энтропии в ней
не приобретет минимальное значение из всех возможных, то есть, пока величина
диссипативная функции не примет минимальное значение."
Таким образом, второе начало термодинамики для живых организмов можно
представить в следующем виде:
I. 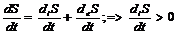 ,
, 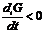
II. 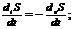
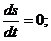 .
. 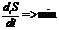
, II, III - второе начало термодинамики для живых организмов.
11. Кинетика биофизических процессов. Уравнение переноса
Термодинамический подход оказывается плодотворным и при описании
кинетических процессов, то есть, скорости их протекания в различных условиях.
Термодинамический анализ кинетики дает возможность рассматривать процессы
разной природы с единой точки зрения, не вдаваясь в частные механизмы. Это
является особенно важным в тех случаях, когда механизм явления очень сложен или
вообще непонятен.
При описании любого явления переноса обобщенной координатой 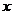 , скорость процесса выражается в виде
ее производной по времени
, скорость процесса выражается в виде
ее производной по времени 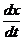 . Во многих случаях скорости процессов прямо пропорциональны
соответствующим обобщенным силам -
. Во многих случаях скорости процессов прямо пропорциональны
соответствующим обобщенным силам -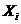 , которая характеризует причины возникновения
соответствующего процесса, тогда это можно представить в следующем виде:
, которая характеризует причины возникновения
соответствующего процесса, тогда это можно представить в следующем виде:
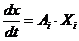
Рассмотрим примеры:
В процессе теплопроводности обобщенной координатой является тепло, а
обобщенной силой - градиент температуры:
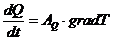 ,
, 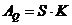
- площадь теплообмена
- коэффициент теплопроводности
Химические реакции:
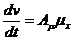
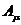 - химическое сродство,
- химическое сродство, 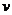 -число молей вещества, вступивших в
реакцию за еденицу времени.
-число молей вещества, вступивших в
реакцию за еденицу времени.
- химический потенциал.
Перенос заряда:
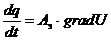 , - закон Ома в дифференциальной форме.
, - закон Ома в дифференциальной форме.
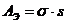 ;
; 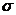 -удельная электропроводность;
-удельная электропроводность; 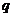 - переносимый заряд; U- разность
потенициалов.
- переносимый заряд; U- разность
потенициалов.
Диффузионные:
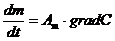 ,
, 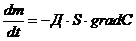
- масса переносимого вещества;
 - коэффициент диффузии;
- коэффициент диффузии;
- площадь, через которую идет данный процесс;
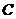 -концентрация.
-концентрация.
В реальных системах практически не бывает изолированных процессов. Они
всегда протекают во взаимосвязи, в частности, между одновременно идущими
процессами происходит обмен энергией. Такие процессы называют термодинамически
сопряженными.
Термодинамическое сопряжение существенно влияет на кинетику биофизических
и биохимических процессов, в результате сопряжения, скорость каждого из них
будет зависеть не только от своей обобщающей силы, но и от всех обобщающих сил,
действующих в системе. С учетом этого, уравнение переноса в общем виде
записывается следующим образом:

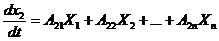
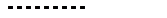
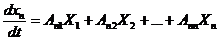
Рассмотрим примеры:
Если в системе (например, в газе) поддерживается одновременно градиент
температуры и градиент концентрации, то уравнение переноса записывается в
следующем виде:
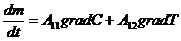
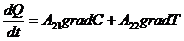
градиент и градиент 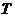 - в данном случае являются обобщающими силами.
- в данном случае являются обобщающими силами.
Если в какой-то части системы и вещества и выше, чем в другой, то градиент и градиент имеют одинаковые знаки, и процессы,
происходящие под действием этих градиентов, усиливают друг друга, то есть масса
переноса больше, чем в условиях существования только одного градиента. Если же
градиенты и - разнонаправлены, то есть, имеют разные знаки, и при этом 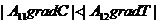 , то
, то 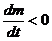 Это означает, что перенос вещества
осуществляется против градиента .
Это означает, что перенос вещества
осуществляется против градиента .
Транспорт веществ, через клеточные мембраны осуществляется благодаря
термодинамическому сопряжению различных процессов, только в тех случаях, когда
молекулы транспортного вещества не имеют заряда, химически и осмотически
инертны, то процесс массы переноса сводится к простой диффузии (закон Фика):
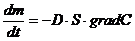
Фактически, такими свойствами в организме обладают только азот и инертные
газы, и близко к этому кислород и углекислота. Во всех остальных случаях,
например, при транспорте воды, солей, углеводов, жирных кислот и т. д., вместе
с концентрационным градиентом действуют и другие градиенты. При транспорте
ионов (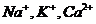 ) по меньшей мере, присутствует три
процесса:
) по меньшей мере, присутствует три
процесса:
. Диффузия вещества.
. Перенос заряда.
. Химическая реакция (в частности, перенос фосфатной группы с АТФ).
В этой связи, уравнение переноса имеет следующий вид:
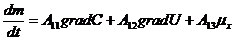
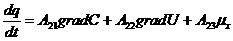
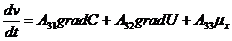
Обобщающими координатами в данной системе уравнений являются m, q и v число
молей АТФ, которая обеспечивает энергией активный транспорт ионов, обобщенными
силами являются: градиент , напряженность электрического поля, химических потенциал
(АТФ).
Рассчитав коэффициент 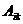 или определив их экспериментально, можно точно знать, в
каком направлении идут кинетические процессы по перемещению ионов через
биологическую мембрану (БМ).
или определив их экспериментально, можно точно знать, в
каком направлении идут кинетические процессы по перемещению ионов через
биологическую мембрану (БМ).
Необходимо отметить, что все, выше рассмотренные уравнения, достаточно
хорошо описывают процессы, которые не так далеко удалены от состояния
равновесия. Так как все уравнения - линейные. Однако, при рассмотрении
процессов, происходящих на БМ, приходится иметь дело с большими градиентами,
что приводит к большим скоростям процессов. Для описания таких процессов
желательно использовать нелинейную термодинамику.
. Кинетика процессов, идущих с преодолением потенциального барьера
Многие реальные процессы в биологической системе идут с преодолением
потенциального барьера. Это означает, что начальное и конечное состояние
системы разделены некоторым промежуточным состоянием, которое обладает большой
энергией.
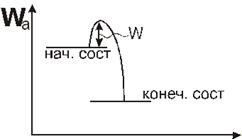
Чтобы процесс осуществлялся, система в начале должна получить
дополнительно энергию 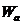 для преодоления потенциального барьера. Величина этой энергии
называется высотой потенциального барьера или энергией активации процесса.
для преодоления потенциального барьера. Величина этой энергии
называется высотой потенциального барьера или энергией активации процесса.
Примером процессов, идущих с преодолением потенциального барьера (ПБ),
служат химические и биохимические реакции. Практически все они проходят стадию
промежуточного состояния с повышенной энергией. В химии такое состояние
называют активированным комплексом.
В реальных телах различные частицы (молекулы, ионы и т. д.) имеют разную
энергию. Поэтому в каждом конкретном случае одни частицы способны преодолевать
ПБ, а другие - нет. В этой связи скорость процессов, в которых участвуют
молекулы данного вещества, будет определяться только числом молекул, которые
способны преодолевать ПБ. Такие частицы (молекулы) называются активными.
Концентрацию активных частиц можно рассчитать по формуле Больцмана:
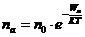
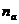 - число активированных частиц;
- число активированных частиц;
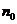 - общее число частиц;
- общее число частиц;
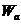 - энергия активации для одного моля вещества.
- энергия активации для одного моля вещества.
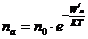
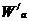 - энергия активации для 1 частицы;
- энергия активации для 1 частицы;
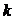 - постоянная Больцмана.
- постоянная Больцмана.
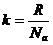 ,
,
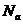 - число Авогадро.
- число Авогадро.
Анализ уравнений Больцмана показывает, что скорость процессов, идущих с
преодолением потенциального барьера (ПБ) экспоненциально зависит от
температуры. Таким образом, зависят скорости испарения, диффузии, химической
реакции и т. д. от температуры. Это имеет важное значение для
жизнедеятельности. В этой связи, температурный гомеостазис организма
гомойотерных животных является необходимым условием стабильной
жизнедеятельности. В противном случае, даже при небольшом отклонении
температуры тела от нормы, скорости различных жизненных процессов изменились бы
значительно, причем, в неодинаковой степени вследствие различия их энергии
активации. Это неизбежно привело бы к нарушению основных функций организма.
Скорость многих химических реакций также определяется энтропией
активации.
Например, при столкновении двух сложных молекул, которые в принципе
взаимодействуют между собой, реакция фактически происходит не всегда. Дело в
том, что молекулы взаимодействуют не целиком, а определенными функциональными
группами. В этом случае можно сказать, что скорость реакции будет
пропорциональна как числу активных частиц, так и вероятности удачных
столкновений.
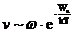
Понятие вероятности в молекулярном движении связано с понятием энтропии:
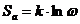
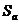 - энтропия активации процесса, следовательно, выражение для
скорости процесса примет вид:
- энтропия активации процесса, следовательно, выражение для
скорости процесса примет вид:
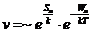
Скорость реакции можно значительно повысить, если каким-то образом
увеличить вероятность удачных столкновений. Для этого необходимо направить
навстречу друг другу соответствующие функциональные группы. Именно так
действуют ферменты. Ферменты - это биологические катализаторы. Роль ферментов
состоит в резком ускорении биохимических реакций. В действии ферментов выделяют
два механизма: одни ферменты в основном снижают энергию активации (); другие увеличивают энтропию
активации .
13.
Квантовомеханические основы биоэенергетики. Основные понятия квантовой механики
Успехи последних десятилетий в изучении живого организма на молекулярном
и субмолекулярном уровнях, позволили проникнуть в молекулярные механизмы его
строения и энергетического обеспечения. Квантовая механика представляет собой
раздел физики (математики), описывающий общие закономерности движения и
взаимодействия микрочастиц. В классической механике четко разграничивают два
вида движения:
. Корпускулярное.
. Волновое
Для корпускулярного движения характерно то, что объект движется по вполне
определенной траектории, и в каждый конкретный момент времени имеет четкую
локализацию в пространстве. Для волнового движения, наоборот, характерна
делокализация в пространстве. Применительно к волне нельзя сказать, что она
находится в данной точке пространства, и не имеет смысла говорить о траектории
волны. Применительно к микрочастицам применяют диалектический дуализм,
рассматривая их и как частицы, и как волны, то есть, имеет место
корпускулярно-волновой дуализм (волны де Бойля). В квантовой механике,
уравнение Шредингера играет важную роль, как и второй закон Ньютона в
классической механике, и уравнение Максвелла в электродинамике. Уравнение
Шредингера в квантовой механике является исходным, основополагающим. И пока оно
не выведено из других соотношений. Справедливость этого уравнения доказывается
только тем, что выводы, полученные с помощью его, находятся в хорошем
соответствии с экспериментальными результатами.
Решения уравнения Шредингера показали, что только свободно движущиеся
частицы могут иметь любую энергию. Что же касается связанных частиц, то есть,
частиц, движущихся в ограниченном пространстве (в пределах атома или молекулы),
то решение уравнения Шредингера для этого случая возможно только при некоторых
определенных значениях энергии. Это означает, что связанная частица может иметь
только дискретные значения энергии, которые называются собственными. В этой
связи электроны атомов характеризуются комбинацией квантовых чисел: 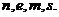
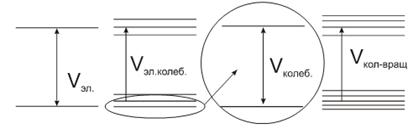
Схема уровней энергии молекул является гораздо более сложной, чем у
атомов. Это связано с тем, что в молекулах возможны, помимо движения электронов
вокруг ядра, также колебательные и вращательные движения.
При колебательных движениях периодически изменяются относительные расположения
ядер в молекуле.
При вращательных движениях изменяется положение в пространстве молекулы,
как целого.
Полная энергия молекулы складывается из трех частей:
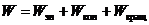
Вклад каждого вида движения в полную энергию не одинаков, то есть:
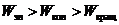
Необходимо заметить, что колебательное и вращательное 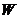 также квантуется.
также квантуется.
При поглощении молекулой энергии могут изменяться все виды энергии,
поэтому полное изменение энергии будет иметь вид:
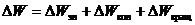
Вклад каждой составляющей - неодинаков:
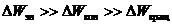
Системы энергетических уровней молекулы представляют собой совокупность
далеко отстоящих друг от друга электронных уровней, при этом каждому
электронному уровню соответствует набор близкорасположенных колебательных
уровней, а каждому колебательному уровню соответствует совокупность еще ближе
расположенных вращательных уровней. Упрощенно графически это можно представить
так:
Изменение электрической энергии 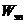 связано с поглощением или с
испусканием кванта энергии, видимой или ультрафиолетовой (УФ) области спектра.
связано с поглощением или с
испусканием кванта энергии, видимой или ультрафиолетовой (УФ) области спектра.
Наряду с электронной энергией при этом процессе могут изменяться 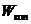 и
и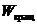 . Поэтому, данному электронному
переходу в спектре соответствует не одна линия, а ряд близко расположенных
линий, которые образуют полосу. В случае простых молекул при наблюдении спектра
приборами большой разрешающей силы, видны линии, составляющие полосу. В сложных
молекулах обычно наблюдается несколько довольно широких полос. Такие спектры
называют электронно-колебательно-вращательными. Они характеризуют молекулу в
целом, и их используют для идентификации вещества.
. Поэтому, данному электронному
переходу в спектре соответствует не одна линия, а ряд близко расположенных
линий, которые образуют полосу. В случае простых молекул при наблюдении спектра
приборами большой разрешающей силы, видны линии, составляющие полосу. В сложных
молекулах обычно наблюдается несколько довольно широких полос. Такие спектры
называют электронно-колебательно-вращательными. Они характеризуют молекулу в
целом, и их используют для идентификации вещества.
При поглощении молекулой не большой порции энергии, не изменяется, тогда, как и могут возрасти. Колебательным
переходам соответствует поглощение в близкой ИК (инфракрасной) области спектра
( ). При данном изменении колебательной
энергии получается полоса, характеризующая колебательно-вращательный спектр.
Эти спектры широко используются для изучения сложных молекул. Многие группы,
входящие в сложные молекулы, характеризуются вполне определенными частотами
колебательных переходов. Эти переходы имеют место при поглощении энергии в
далекой ИК области спектра (
). При данном изменении колебательной
энергии получается полоса, характеризующая колебательно-вращательный спектр.
Эти спектры широко используются для изучения сложных молекул. Многие группы,
входящие в сложные молекулы, характеризуются вполне определенными частотами
колебательных переходов. Эти переходы имеют место при поглощении энергии в
далекой ИК области спектра ( ). Изучая все эти спектры, можно получить достоверную
информацию о строении сложных молекул. А это, в свое время, позволяет по
базовым данным осуществить из идентификацию.
). Изучая все эти спектры, можно получить достоверную
информацию о строении сложных молекул. А это, в свое время, позволяет по
базовым данным осуществить из идентификацию.
14.
Квантовомеханические особенности строения биомолекул
Живые системы на 99% состоят из атомов 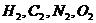 . Большую роль биохимических
процессов играют атомы
. Большую роль биохимических
процессов играют атомы 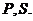 Для атомов
Для атомов 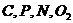 и характерно наличие не поделенной пары электронов и
способность образовывать кратные связи. Состояние электронов в атоме
описывается волновыми функциями АО. Эти атомные орбиты (АО) характеризуют
движение электронов вокруг ядра. В зависимости от орбитального квантового
числа, АО могут быть
и характерно наличие не поделенной пары электронов и
способность образовывать кратные связи. Состояние электронов в атоме
описывается волновыми функциями АО. Эти атомные орбиты (АО) характеризуют
движение электронов вокруг ядра. В зависимости от орбитального квантового
числа, АО могут быть 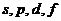 . АО, которые используются для образования связей, называются
валентными орбитами.
. АО, которые используются для образования связей, называются
валентными орбитами.
Состав электронов в молекуле также может быть описан с помощью волновой
функции, которые именуются молекулярными орбитами МО. МО образуются путем
перекрытия атомных орбиталей, в зависимости от того, какие атомные орбитали
образуют данную МО различают и 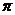 орбитали. Необходимо отметить, что именно электроны находятся
на орбитали обусловливая образование
кратных связей. Для атомов
орбитали. Необходимо отметить, что именно электроны находятся
на орбитали обусловливая образование
кратных связей. Для атомов 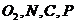 характерно наличие не поделенной пары электронов и
образование кратных связей.
характерно наличие не поделенной пары электронов и
образование кратных связей.
Все многоатомные молекулы могут быть разделены на две группы:
) несопряженные системы;
) сопряженные системы.
Несопряженные системы содержат только одинарные связи или же, в них
имеются изолированные кратные связи, отделенные друг от друга или от атомов с
неподеленной парой электронов, по крайней мере, одним атомом с насыщенной
валентностью. Такие молекулы можно рассматривать, как состоящие из примыкающих
друг к другу и почти независимых друг от друга, связей. Каждая связь описывает
локализованную двуцентровую МО, аналогично молекулярным орбитам в двухатомной
молекуле. Свойства образующих молекулярных связей постоянны и для одной и той
же связи, входящей в разные молекулы, свойства меняются незначительно.
Сопряженные системы представляют собой молекулы, содержащие несколько
кратных связей, в которых участвуют соседние атомы или же молекулы, в состав
которых входят атомы с неподеленными электронными парами, расположенными рядом
с кратной связью. Для таких систем характерно нелокализованные многоцентровые
молекулярные орбитали, которые относятся к молекуле в целом или к большей ее
части.
Если молекула с сопряженными связями, содержит атомы, обладающие
неподеленными электронными парами, то электронами этих пар могут включаться в
общую систему - электронов. Существование делокализованных-электронов является важнейшим
свойством молекул с сопряженными связями. Основные химические,
физико-химические и биохимические свойства таких систем определяются - электронами, так как они более
подвижны, по сравнению с -электронами и с большей легкостью вступают в химические реакции.
15. Спектры
поглощения сложных молекул
Важным
источником информации о структуре сложных молекул являются их спектр и
поглощение. Излучение спектров поглощения в УФ и в видимой областях позволяют
получать информацию о системе электронных энергетических уровней молекулы и
вероятности перехода между этими уровнями. В основном состоянии молекулы все ее
электроны расположены по низшим молекулярным орбиталям, выше которых расположен
ряд свободных орбиталей с более высокой энергией. Низшее возбужденное состояние
соответствует переходу электрона с высшей заполненной на низшую свободную
орбиталь.
Система МО сложных органических и биологически важных молекул состоит из
локализованных - орбиталей, локализованных и делокализованных - орбиталий, а также, 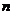 -орбиталей, на которых находятся
электроны неподеленных пар. Заполненные орбитали обозначаются символами , , , а свободные орбитали -
-орбиталей, на которых находятся
электроны неподеленных пар. Заполненные орбитали обозначаются символами , , , а свободные орбитали - 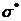 ,
, 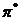 .
.
Порядок возрастания энергии обычно бывает таким:
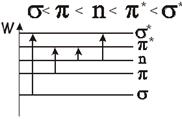
Среди этих переходов, наибольший интерес представляют 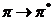 , соответствующие этим переходам
полосы в спектрах поглощения, обычно имеют довольно большую интенсивность в
ближней УФ или видимой области спектра. Полосы, соответствующие
, соответствующие этим переходам
полосы в спектрах поглощения, обычно имеют довольно большую интенсивность в
ближней УФ или видимой области спектра. Полосы, соответствующие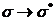 , обычно лежат в далекой УФ части
спектра. При
, обычно лежат в далекой УФ части
спектра. При 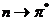 , или
, или 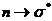 , один из электронов неподеленной пары перемещается соответственно
на или орбиталь. Полосы, соответствующие
этим переходам, имеют крайне малую интенсивность.
, один из электронов неподеленной пары перемещается соответственно
на или орбиталь. Полосы, соответствующие
этим переходам, имеют крайне малую интенсивность.
Для характерно то, что при наличии системы сопряженных связей,
поглощенная энергия кванта света передается не отдельному электрону, а всей
электронной системе, поэтому наличие молекул делокализованных - электронов может быть обнаружено по
присутствию в спектрах поглощения характерных полос.
16. Транспорт
веществ в организме. Биомембранология. Структура и функции биологических
мембран
Для организма, как для открытой системы, характерен обмен с окружающей
средой энергией, веществом и информацией. Необходимым условием обмена веществ
является их транспорт внутри биологической системы. Все вещества, из которых
состоит организм, перемещаются в нем с потоком крови и лимфы, они преодолевают
значительные расстояния, обеспечивая обмен между органами и тканями. В ходе
обмена между кровью и тканями, вещества проникают сквозь стенки кровеносных
сосудов. Внутри клеток происходит непрерывный транспорт веществ через мембраны
органелл. Заболевания, при которых нарушен транспорт веществ в организме,
связаны, прежде всего, с различными нарушениями молекулярной организации
биологических мембран - (БМ).
Термин "мембрана" используют в биологии, обозначая им клеточную
границу, которой свойственна полупроницаемость, то есть, легкость проникновения
сквозь нее одних веществ при невозможности преодоления ее другими.
Каждая клетка окружена наружной мембраной, которая называется
плазматической мембраной или плазмолеммой. В настоящее время клетку стали
воспринимать, как обширную сеть мембранных систем, составляющих важнейший
элемент клеточной организации; соотношение между плазмолеммой и внутриклеточной
мембраной неодинаково в различных клетках. Так, в клетках хрусталика глаза нет
никаких других мембран, кроме плазматических, тогда, как в почечных клетках
(гепатоцитах), площадь плазматических мембран составляет порядка 6% от
поверхности, занимаемой всеми мембранами клетки. Установлено, что по мере
увеличения отношения сумм площади мембран к объему клетки, повышается
интенсивность обменных процессов в этой клетке.
Электронная микроскопия клеточных мембран не позволяет их увидеть, так
как они состоят из тех же химических элементов, что и цитоплазма. Для получения
четкой электронограммы клетки, ее мембраны контрастируют. Для этого производят
осаждение на них вольфрама, осмия и других элементов, которые хорошо поглощают
и рассеивают электроны. На таких препарированных клетках любая БМ выглядит
трехслойной: между двумя темными полосами располагается светлое пространство.
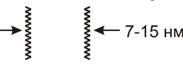
Следовательно, компоненты промежуточной части БМ слабо связывают входящие
в состав красители. Суммарная толщина трехслойной структуры может изменяться от
7 до 15 нм, причем, разная величина присуща различным клеточным мембранам. Во
многих БМ наблюдается асимметрия трехслойной организации, то есть, темные
полосы различны по ширине и по плотности.
Все клеточные мембраны построены в основном из липидов, белков
(протеинов) и углеводов. При этом, углеводы образуют соединения с белками
(гликопротеины) и липидами (гликолипиды). Органические вещества образуют соли с
различными ионами, которые присутствуют в виде водных растворов внутри
мембранных каналов.
Структурной основой БМ служит бимолекулярный слой. Его образование
обусловлено особенностями взаимодействия с водой мембранных липидов, среди
которых преобладают фосфолипиды. В молекуле фосфолипидов условно выделяются 3
части:
головка;
тело;
пара длинных хвостов.
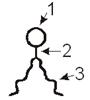
Головка (1) образована одним из таких соединений, как холин, серин,
треонин, инозин, эталомин; Головка содержит и остаток фосфорной кислоты.
Тело (2) состоит из молекулы глицерина или сфингозина.
Головка фосфолипида либо электронейтральна, либо несет отрицательный
заряд. Образующие ее молекула, а также глицерин или сфингозин - гидрофильны,
тогда, как жирно-кислотные хвосты - гидрофобны. В этой связи, в воде молекулы
фосфолипидов самопроизвольно выстраиваются так, что их гидрофобные
углеводородные цепи оказываются укрытыми от воды, а головки вступают во взаимодействие
с водой. В результате создается конструкция, поперечник которой включает две
молекулы фосфолипидов, повернутых друг к другу жирно-кислотными хвостами и
обращенные к обеим наружным поверхностям гидрофильными головками. Графически
это можно представить так:
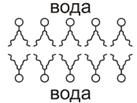
Так образуется сплошной бимолекулярный фосфолипидный слой (бислой),
который и служит каркасом биологической мембраны. Для создания 1 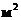 поверхности БМ достаточно 1 мг
фосфолипида. При этом, на каждый квадратный микрон клеточной мембраны,
приходится примерно
поверхности БМ достаточно 1 мг
фосфолипида. При этом, на каждый квадратный микрон клеточной мембраны,
приходится примерно 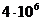 липидных молекул, то есть, по 2 млн в каждом из моно слоев
бесслойной структуры.
липидных молекул, то есть, по 2 млн в каждом из моно слоев
бесслойной структуры.
Наряду с фосфолипидами, в состав биологических мембран могут входить и
другие жиры, причем, характерным свойством большинства из них, является наличие
пары углеводородных хвостов. Одноцепочные липиды встречаются крайне редко. В их
присутствии, клеточные мембраны имеют тенденцию к разрушению. С этим связана причина
гибели людей при укусах тех видов змей, в яде которых имеется особый фермент -
фосфолипаза 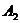 . Он превращает двухцепочные фосфолипиды в одноцепочный липид
- лизолецитин, в результате чего мембраны клеток разрушаются. Другой фермент -
фосфолипаза , выделяемый некоторыми микроорганизмами (например, холерным
виброном), разрушает БМ, "откусывая" полярные головки фосфолипидов.
. Он превращает двухцепочные фосфолипиды в одноцепочный липид
- лизолецитин, в результате чего мембраны клеток разрушаются. Другой фермент -
фосфолипаза , выделяемый некоторыми микроорганизмами (например, холерным
виброном), разрушает БМ, "откусывая" полярные головки фосфолипидов.
Особое место среди мембран липидов, занимает холестерин. По мере
повышения содержания холестерина в БМ, площадь, занимаемая фосфолипидами
сокращается до тех пор, пока на одну молекулу холестерина будет приходиться две
молекулы фосфолипида. Эффект сокращения площади, приходящийся на одну молекулу
фосфолипида, обусловлен тем, что изменяется наклон его углеводородных цепей к
поверхности бислоя. По мере увеличения содержания холестерина в БМ, фосфолипиды
стремятся встать перпендикулярно мембранной поверхности. В результате чего,
укладка фосфолипидов приобретает большую компактность, и мембрана уплотняется.
В результате, она становится более вязкой и менее проницаемой для многих
веществ (например, глюкозы, ионов воды и т. д.). Содержание холестерина в
клетках зависит от общего холестеринового обмена в организме, который в свою
очередь сильно подвержен влиянию пищевого рациона.
17.
Мембранные белки
В липидный каркас клеточных мембран встроены белковые компоненты
(протеины). На каждую клетку в среднем приходится около 10 пг мембранных белков
(МБ). Различают периферические и собственные (интегральные) белки биологических
мембран. Белки первого типа располагаются на поверхности липидного бислоя.
Здесь связь между липидами и белками осуществляется за счет электростатического
взаимодействия между противоположно заряженными полярными группами этих
веществ. Интегральные белки взаимодействуют своими гидрофобными областями с
углеводородными цепочками липидов только за счет Ван-дер-ваальсовых сил. У
интегральных белков все гидрофильные области спрятаны внутрь молекулы, а наружу
направлены неполярные группы. В этой связи, собственные белки полностью или
частично погружаются в биологическую мембрану, при этом крупные белковые
молекулы могут пронизывать БМ насквозь.
Наиболее характерным типом вторичных структур интегральных белков
является спираль. Причем, на участках, окруженных липидами, преобладает 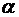 -спираль, у которой все
аминокислотные остатки - гидрофобные. Реже встречается
-спираль, у которой все
аминокислотные остатки - гидрофобные. Реже встречается 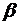 - спираль, представляющая собой полое
микро трубчатое образование, наружные стенки которого сосредоточены гидрофобные
аминокислотные остатки, а внутрь полости ориентированы гидрофильные группы.
Предполагается, что белки, имеющие структуру - спирали, образуют ионные каналы в
клеточных мембранах.
- спираль, представляющая собой полое
микро трубчатое образование, наружные стенки которого сосредоточены гидрофобные
аминокислотные остатки, а внутрь полости ориентированы гидрофильные группы.
Предполагается, что белки, имеющие структуру - спирали, образуют ионные каналы в
клеточных мембранах.
Функции мембранных белков весьма разнообразны. Среди них можно выделить
следующие основные:
. Обеспечение транспортных свойств БМ (образование мембранных каналов,
работа в качестве переносчиков определенных веществ, участие в системах
активного транспорта);
. Белки несут функцию структурного компонента БМ, тем самым, они
усиливают прочность липидного каркаса.
. Большим разнообразием различаются мембранные протеины, катализирующие
химические реакции. Причем, белки-ферменты, встроенные в БМ, действуют
активнее, чем при их пребывании в растворе. Установлено, что основная часть
всех биохимических реакций, протекает на клеточных мембранах.
. Важным функциональным типом МБ являются белки-рецепторы. Они столь
разнообразны, сколь разнообразны стимулы, которые воспринимаются рецепторными
белками. Так, например, восприятие света, обеспечивается пигментами, в состав
которых, наряду с белками, входят хромофорные группы с определенным спектром
поглощения. Таким пигментом является родопсин, который обеспечивает реакции на
свет палочек или колбочек.
. Белки, присутствующие в мембранах любой клетки, осуществляют ее
взаимодействие с окружающей средой, а также обмен информацией между органеллами
внутри клетки.
. Белки, образующие антитела, способны связывать специфический антиген
(белок бактерии, вируса, токсина и т. д.) и вызывать иммунный ответ клетки, в
плазмолемме которой белки сосредоточены.
В различных мембранах белки распределены среди липидов по-разному. В
плазмолемме их распределение довольно равномерное. Для специализированных
внутриклеточных мембран, характерно неоднородное распределение белков в БМ.
Различия между клеточными мембранами разных типов, между участками БМ и
даже, между сторонами одной и той же мембраны, привели к выводу о гетерогенности
БМ.
Под гетерогенностью понимают разнородность их структур и функциональных
свойств. Особенности молекулярных структур клеточных мембран и предопределяют
их физические и физико-химические свойства.
18. Физические и физико-химические свойства БМ
В живых клетках БМ представляют собой жидкокристаллические структуры.
Структуры жидкого кристалла образуют как липидные, так и белковые молекулы.
Молекулярной организации клеточных мембран, как жидкого кристалла, характерно
состояние с высокой степенью упорядоченности и текучести (лабильности).
БМ присущи такие свойства:
. Значительная прочность на разрыв.
. Упругость (эластичность).
. Поверхностное натяжение.
. Вязкость.
. Электрострикция.
. Флексоэлектрострикция.
Два последних свойства обусловлены наличием повторного заряда на БМ.
. Поверхностный заряд на клеточной мембране (КМ)
Он создается полярными головками фосфолипидов, гликопродеидами (главным
образом, карбоксильными группами сиаловой кислоты и аминокислотными остатками),
а также, гликолипидами. За счет этих веществ, поверхность БМ заряжена
отрицательно. Поверхностный заряд плазмолеммы играет важную роль в межклеточных
взаимодействиях. Он способствует стабильности мембранных структур, а также,
связыванию ионов, находящихся в межклеточной среде. От поверхностного заряда
плазмолеммы зависит ионный состав предмембранных слоев, что оказывает влияние
на многие внутриклеточные обменные процессы.
Существование заряженных групп на БМ приводит к образованию двойного
диффузионного электронного слоя, в котором фиксированный отрицательный заряд
клеточной поверхности уравновешен положительным зарядом, который создается
межклеточной средой за счет положительных ионов. Подвижность ионов не одинакова
в различных условиях. Разность потенциалов между неподвижными и подвижными
частями двойного электронного слоя называют электрокинетическим потенциалом или
дзета потенциалом. Толщина двойного электрического слоя и дзета потенциала
зависит от природы электролита и концентрации ионов (в межклеточной среде).
Так, при уменьшении концентрации 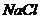 в межклеточной среде в 200 раз,
толщина двойного электрического слоя возрастает в 5 раз. При высокой
концентрации электролита, дзета потенциал стремиться к нулю. Когда в
межклеточной среде присутствуют двухвалентные катионы, избыток положительного
заряда может стать настолько велик, что дзета потенциал изменит свой знак.
Снижение дзета потенциала и, тем более, изменение его знака на противоположный,
вызывает слипание плазматических мембран соседних клеток. Это проявляется при
избытке ионов в межклеточной среде, а также, при перестройках клеточных
мембран, приводящих к сдвигам дзета потенциала. Подобный эффект имеет свои
особенности у клеток крови, в которых дзета потенциал форменных элементов
(эритроцитов) может падать за счет нарушения не только солевого, но и белкового
состава кровяной плазмы. Это явление лежит в основе изменения скорости
осаждения эритроцитов (СОЭ), измерение которых широко используется в
диагностических целях.
в межклеточной среде в 200 раз,
толщина двойного электрического слоя возрастает в 5 раз. При высокой
концентрации электролита, дзета потенциал стремиться к нулю. Когда в
межклеточной среде присутствуют двухвалентные катионы, избыток положительного
заряда может стать настолько велик, что дзета потенциал изменит свой знак.
Снижение дзета потенциала и, тем более, изменение его знака на противоположный,
вызывает слипание плазматических мембран соседних клеток. Это проявляется при
избытке ионов в межклеточной среде, а также, при перестройках клеточных
мембран, приводящих к сдвигам дзета потенциала. Подобный эффект имеет свои
особенности у клеток крови, в которых дзета потенциал форменных элементов
(эритроцитов) может падать за счет нарушения не только солевого, но и белкового
состава кровяной плазмы. Это явление лежит в основе изменения скорости
осаждения эритроцитов (СОЭ), измерение которых широко используется в
диагностических целях.
Дзета потенциал, измеряемый у различных клеток, изменяется в пределах от
-10 до -30 мВ. Его величина спадает по exp закону, с увеличением расстояния от
поверхности БМ. Дзета потенциал оказывает существенное влияние на размеры
межклеточных пространств, и противодействует силам их притяжения. Наименьшее
расстояние между клетками составляет порядка 10-20 нм. При таком расстоянии
существует энергетическая "яма" во взаимодействии кулоновских и
Ван-дер-Ваальсовских сил. В большинстве тканей расстояние между клетками составляет
от 0,1 до 1 микрона.
Липидная часть БМ обладает свойствами диэлектрика. Относительная
диэлектрическая проницаемость ( ) гидрофобной зоны составляет порядка 2-2,2, тогда, как в
гидрофильной части гораздо выше, и составляет около 10-20. Емкость полярных
головок липидов достигает примерно 30
) гидрофобной зоны составляет порядка 2-2,2, тогда, как в
гидрофильной части гораздо выше, и составляет около 10-20. Емкость полярных
головок липидов достигает примерно 30 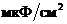 , а жирно-кислотные хвосты имеют
емкость около 0,5-0,9 . Общая емкость БМ составляет примерно 1. Благодаря существованию в БМ
заряженных групп, ей присуща ярко выраженная электрострикция, то есть, по мере
повышения трансмембранной разности потенциалов, БМ сжимается, что приводит к
утоньшению гидрофобной зоны мембраны и, соответственно, к увеличению мембранной
емкости. Возрастание дзета потенциала сопровождается увеличением клеточной
поверхности.
, а жирно-кислотные хвосты имеют
емкость около 0,5-0,9 . Общая емкость БМ составляет примерно 1. Благодаря существованию в БМ
заряженных групп, ей присуща ярко выраженная электрострикция, то есть, по мере
повышения трансмембранной разности потенциалов, БМ сжимается, что приводит к
утоньшению гидрофобной зоны мембраны и, соответственно, к увеличению мембранной
емкости. Возрастание дзета потенциала сопровождается увеличением клеточной
поверхности.
. Механические свойства БМ
Механические свойства КМ весьма своеобразны. Величина модуля Юнга в
поперечном направлении достигает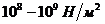 . Вместе с тем, устойчивость к деформации сдвига примерно на
4 порядка меньше. Сила, достаточная для разрушения БМ на участке в 1 мкм,
оценивается величиной примерно
. Вместе с тем, устойчивость к деформации сдвига примерно на
4 порядка меньше. Сила, достаточная для разрушения БМ на участке в 1 мкм,
оценивается величиной примерно 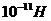 . Значительный вклад в повышение упругости и прочности
мембраны вносят не только интегральные, но, также, довольно густая цепь
периферических белков.
. Значительный вклад в повышение упругости и прочности
мембраны вносят не только интегральные, но, также, довольно густая цепь
периферических белков.
21. Вязкость БМ
В живой клетке БМ находятся в жидкокристаллическом состоянии.
Жидкокристаллическому состоянию присущи текучесть и определенная вязкость.
Величина вязкости была измерена с помощью ЭПР (эл. парамагнитного резонанса) и
ДСК (диффузионно - сканирующей микрокаллометрией). Результаты измерения
вязкости разными методами совпали. При этом вязкость БМ изменилась от 0,03 до
0,1 Па*с (30-100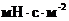 ) или 30-100 сП (сантипуаз). Это примерно в 30-100 раз
больше, чем вязкость воды и примерно такая же, как вязкость оливкового или
подсолнечного масла. При изменении температуры молекулярного состава и других
параметров биологических мембран, изменяется ее вязкость, вследствие чего
происходят фазовые изменения в структуре мембраны, то есть, осуществляется
переход жидкий кристалл - твердый кристалл.
) или 30-100 сП (сантипуаз). Это примерно в 30-100 раз
больше, чем вязкость воды и примерно такая же, как вязкость оливкового или
подсолнечного масла. При изменении температуры молекулярного состава и других
параметров биологических мембран, изменяется ее вязкость, вследствие чего
происходят фазовые изменения в структуре мембраны, то есть, осуществляется
переход жидкий кристалл - твердый кристалл.
Одним из важных достоинств температурного гомеостаза является поддержание
БМ в жидкокристаллическом состоянии, что обеспечивает стабильность транспорта
веществ через нее. Жизнеспособность организма страдает как при понижении, так и
при повышении мембранной проницаемости, которая, в свою очередь, зависит от
вязкости БМ.
22.
Подвижность молекулярных компонентов БМ
Основными формами молекулярного движения в БМ являются:
. Латеральная миграция (перемещение молекул в плоскости мембраны в
пределах одной стороны бимолекулярного слоя).
. Вращение молекул вокруг собственной оси.
Скорости обоих процессов примерно такие же, как скорость свободной
диффузии в вязкой среде.
Большой свободой движения обладают липиды. Среднее время пребывания
фосфолипидной молекулы в данной точке мембраны не более 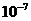 с, следовательно, мембранные липиды
за счет латеральной миграции постоянно меняются местами, причем, каждая
молекула меняет своих соседей примерно
с, следовательно, мембранные липиды
за счет латеральной миграции постоянно меняются местами, причем, каждая
молекула меняет своих соседей примерно 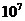 раз в секунду, передвигаясь со
скоростью 5-10 мкм в секунду. В жидкокристаллической структуре молекулы
перемещаются скачками. Между частотой перескоков, площадью, занимаемой
молекулой в БМ и средним расстоянием, проходимым молекулой за время t,
установлены следующие соотношения:
раз в секунду, передвигаясь со
скоростью 5-10 мкм в секунду. В жидкокристаллической структуре молекулы
перемещаются скачками. Между частотой перескоков, площадью, занимаемой
молекулой в БМ и средним расстоянием, проходимым молекулой за время t,
установлены следующие соотношения:
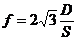 - частота перескоков;
- частота перескоков;
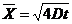 - среднее расстояние;
- среднее расстояние;
- площадь, занимаемая одной молекулой;
- коэффициент диффузии или коэффициент латеральной миграции.
Его величина у мембран фосфолипидов обычно составляет 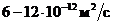
Скорость вращательного движения мембран молекул довольно велика, и она
зависит от размеров молекулы и свойств мембраны. Так, поворот на 1 рад
фосфолипид совершает примерно за 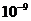 с; родопсин - за
с; родопсин - за 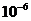 с; цитохромоксидаза - фермент
дыхательных цепей митохондрий - за
с; цитохромоксидаза - фермент
дыхательных цепей митохондрий - за 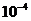 с.
с.
В отличие от вращения и лотеральной миграции, перемещение молекул поперек
мембраны, то есть, с одной стороны липидного слоя на другой, совершаются очень
редко.
Асимметрия клеточных мембран имеет важное значение в переносе через нее
различных веществ. Асимметрия строения белков и липидов БМ обеспечивает
векторный транспорт через БМ, то есть, однонаправленный перенос веществ через
нее.
Лабильность БМ проявляется не только в значительной подвижности их
молекулярных компонентов, но и в высоких темпах обновления. Темп обновления
компонентов оценивается временем полу жизни каждого из них. Средние показания
этого времени различны у белков в разных мембранах. Так, в плазмолемме и
мембранах эндоплазматической сети, оно составляет примерно 50 часов. В
мембранах митохондрий - примерно 120 часов. Липиды также обновляются довольно
быстрыми темпами. У разных фосфолипидов время полу жизни примерно от 15 до 80
часов, у холестерина - от 24 до 140 часов, следовательно, в течение жизни
клетки, ее мембранные компоненты многократно обновляются. Однако, это не
приводит к замене целых мембранных систем, так как скорости обновления молекул
различных типов в одних и тех же биологических мембранах меняются в широких
пределах.
23. Функции
биологических мембран
Физические, химические и физико-химические свойства биологических мембран
предопределяют выполнение ими определенных функций, без которых
жизнедеятельность организма невозможна. В самом общем виде, и применительно ко
всем мембранам, выделяют три основные функции БМ:
. Барьерная.
. Матричная.
. Механическая.
Механическая функция заключается в поддержании морфологической
целостности и относительной автономности как клетки в целом, так и
внутриклеточных органелл. Эта функция основана, прежде всего на механических
свойствах мембранных структур.
Под барьерной функцией понимается создание биомембранных препятствий для
свободного перемещения веществ через нее. Для одних веществ мембраны являются
непреодолимым препятствием, другие легко проходят сквозь нее, причем, как
правило, только в одном направлении. Скорость мембранного переноса разных
веществ неодинакова. Следовательно, с барьерной функцией БМ непосредственно
связана ее избирательная селективная проницаемость.
Мембраны не только отделяют друг от друга клетки, но, также, разделяют
цитоплазму на ряд замкнутых отсеков ("компартментов"), каждый из
которых выполняет свою специфическую функцию. В такой трактовке понятие
компартментов совпадает с понятием органелл. Благодаря компартментализации, в
клетке пространственно разобщены, и изолированы друг от друга биохимические
процессы, течение которых совместно невозможно. Так, например, синтез жирных
кислот происходит в цитоплазме, а их окисление - в митохондриях. Синтез белка
происходит на рибосомах, а его деградация - в лизосомах. Между содержимым
органелл и цитозолем имеются существенные различия в химическом составе, что
обусловливает наличие высоких концентрационных градиентов на внутриклеточных
мембранах. На плазмолемме также поддерживаются значительные физико-химические
градиенты. Градиенты служат главной движущей силой трансмембранного переноса
веществ.
Преимущества компартментализации связаны не только с барьерной, но и с
матричной функцией клеточных мембран. БМ служит матрицей для белков-рецепторов,
ферментов и других физиологически активных веществ. Матричная функция
обеспечивает пространственную организацию рецепторных взаимодействий
метаболических реакций, переноса энергии и других мембранных процессов. Так, в
БМ объединяются встроенные в них ферменты в единый конвейер, где каждый из них
действует строго согласовано с остальными. Среди мембранных ферментов
выделяются векторные, которые пронизывают БМ насквозь и обеспечивают прием
одних компонентов на ее одной стороне, и выделение продуктов реакции на
противоположной стороне.
Различные клеточные мембраны осуществляют также ряд специфических,
присущих только им, функций. К характерным функциям плазмолеммы относят:
. Механическая защита клетки.
. Обеспечение межклеточных взаимодействий.
. Электрическая изоляция клетки.
. Экранировка клетки от внешних электромагнитных полей.
. Биоэлектрогенез.
. Генерация нервных импульсов (потенциалодействие) и их проведение по
нервным волокнам.
24.
Биофизические механизмы тарнспорта веществ через БМ
Различают пассивный и активный транспорт веществ через клеточные
мембраны. К пассивному относится транспортно-мембранный массоперенос,
происходящий в направлении действия концентрационного, электрического,
осмотического, фильтрационного (гидростатического) градиентов. Так как, по
отношению к одному и тому же веществу перечисленные градиенты могут быть
направлены противоположно, то при анализе массопереноса необходимо учитывать
термодинамическое сопряжение всех физико-химических градиентов.
Активным транспортом называют перенос вещества в направлении,
противоположном тому, которое предопределено термодинамическим сопряжением всех
перечисленных выше градиентов. Движущей силой активного транспорта служит
химический потенциал, который обусловливает в БМ течение ферментативных реакций
и поставляет свою энергию, необходимую для преодоления градиентов.
Концентрационный, электрический, осмотический и фильтрационный градиенты
имеют неодинаковое значение в переносе конкретных веществ через клеточные
мембраны. Так, массоперенос, происходящий в результате суммарного действия
осмотического и фильтрационного градиентов, принято называть конвекционным или
конвективным потоком. Этим потоком определяется, прежде всего,
транспортно-мембранный перенос воды вместе с растворенными в ней веществами.
Конвекционный поток обеспечивает обмен веществ в капиллярах между кровью и
межклеточной средой, а также образование первичной мочи в почках.
Через клеточные мембраны пассивный транспорт заряженных частиц (ионов)
происходит главным образом, под воздействием концентрационного и электрического
градиентов, так как, между цитоплазмой и межклеточной средой нет существенных
отличий в осмотическом и гидростатическом давлениях, а, следовательно,
отсутствуют соответствующие градиенты. В этом случае уравнение для пассивного
транспорта ионов через БМ можно представить в следующем виде:
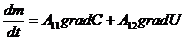
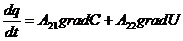
Решение этих двух уравнений приводит к электродиффузионному уравнению
Нернста -Планка, и поток определяется по формуле:
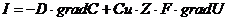
- коэффициент диффузии  ;
;
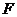 - число Фарадея;
- число Фарадея;
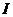 - плотность потока вещества (ионов) через БМ
- плотность потока вещества (ионов) через БМ 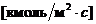 ;
;
- концентрация  ;
;
- подвижность заряженных частиц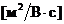 ;
;
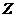 - валентность переносимых ионов;
- валентность переносимых ионов;
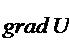 - напряженность электрического поля.
- напряженность электрического поля.
Суммарный массоперенос, обусловленный концентрационным и электрическим
градиентами, может быть, как больше, так и меньше, по сравнению с изолированным
действием каждого из градиентов. В первом случае, градиент направлен в одну
сторону, во втором - в противоположную.
Транспортно-мембранный перенос незаряженных частиц подчиняется уравнению
Фика, которое имеет вид:
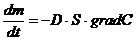
- площадь, через которую осуществляется перенос.
При наличии только градиента, масса переноса описывается только
этим уравнением. Следует отметить, что свободная диффузия под воздействием
только градиента , характерна для небольшого количества веществ в организме (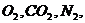 неполярные жирорастворимые
соединения, вирусы, бактерии и т. д.).
неполярные жирорастворимые
соединения, вирусы, бактерии и т. д.).
25.
Коэффициент проницаемости БМ
В биофизике используется понятие коэффициента проницаемости, который
зависит от коэффициента диффузии, от толщины БМ и коэффициента распределения
вещества между липидной частью мембраны и водными фазами, окружающими мембрану.
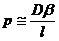
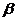 - коэффициент распределения;
- коэффициент распределения;
- толщина мембраны.
Под проницаемостью понимают способность БМ пропускать сквозь себя
определенные вещества, при этом следует помнить, что нет понятия проницаемости
вообще, а есть разная проницаемость мембран для тех или иных веществ.
Оценить мембранную проницаемость для веществ разной природы, можно на
основе анализа коэффициента , через липидный бимолекулярный слой БМ легко проникают те
вещества, у которых достаточно велик. рассчитывается по формуле:
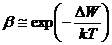
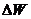 - разность энергий, которой обладает данная частица в воде и
в липиде. Она складывается из двух компонентов:
- разность энергий, которой обладает данная частица в воде и
в липиде. Она складывается из двух компонентов:
) разности энергии гидрофобного взаимодействия;
) разности электронной энергии, характеризующей притяжение заряженной
частицы диполями воды с одной стороны и липидами - с другой.
Переносу ионов из цитозоля или межклеточной среды в БМ свойственно
значительное преобладание второго компонента над первым. Другими словами,
переход ионов из раствора в БМ будет определяться главным образом разностью
электростатических сил (энергии), с которыми на ион действуют диполи воды и
диполи липидов. Мерой взаимодействия ионов с диполями воды и липидов служит
относительная диэлектрическая проницаемость -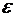 . У воды
. У воды  примерно равна 81, у липидов
примерно равна 81, у липидов 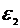 примерно равна 2-3. Электрическую
энергию, которую необходимо преодолеть для переноса одного иона из межклеточной
среды в биомембрану, рассчитывают по формуле Борна, которая имеет вид:
примерно равна 2-3. Электрическую
энергию, которую необходимо преодолеть для переноса одного иона из межклеточной
среды в биомембрану, рассчитывают по формуле Борна, которая имеет вид:
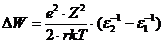
 - заряд электрона;
- заряд электрона;
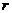 - радиус переносимого иона.
- радиус переносимого иона.
Для переноса одновалентного иона 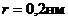 , необходимо затратить энергию,
равную 70 кТ, что соответствует
, необходимо затратить энергию,
равную 70 кТ, что соответствует 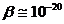 . Такая величина коэффициента означает, что переход иона из
межклеточной водной среды в биологическую мембрану практически невозможен. У
гидрофобных веществ коэффициент на несколько порядков больше, так как они слабо
взаимодействуют с диполями воды и, кроме того, им свойственно гидрофобное
взаимодействие с мембранными липидами, что способствует их переходу в липидную
среду.
. Такая величина коэффициента означает, что переход иона из
межклеточной водной среды в биологическую мембрану практически невозможен. У
гидрофобных веществ коэффициент на несколько порядков больше, так как они слабо
взаимодействуют с диполями воды и, кроме того, им свойственно гидрофобное
взаимодействие с мембранными липидами, что способствует их переходу в липидную
среду.
Следовательно, жирорастворимые вещества способны проникать
непосредственно через липидный каркас клеточной мембраны, растворяясь и
диффундируя в ней. Гидрофильные вещества лишены такой возможности. Для их
трансмембранного переноса используют два основных механизма:
. Ион может образовывать липофильный комплекс с переносчиком
миграционного типа и диффундировать в таком "чехле" сквозь липидный
бислой.
. Можно понизить энергию, затрачиваемую на перенос ионов, за счет
увеличения диэлектрической проницаемости БМ, поскольку в БМ могут присутствовать
сквозные поры (каналы), заполненные водой. В этом случае, внутри каналов такая же, как и у межклеточной
жидкости, и движение ионов по каналам не связано с преодолением высокого
энергетического барьера. Скорость транспорта веществ по каналам на 2-3 порядка
выше, чем при миграции с переносчиком. Переносчик с транспортным веществом
диффундирует в липиде, и его движение сильно зависит от вязкости БМ. Движение
вещества по каналам не зависит от вязкости БМ.
26. Свободная
диффузия жирорастворимых веществ
Установлено, что вещества тем легче проникают в клетку, чем выше их
растворимость в липидах, а она высока у неполярных (гидрофобных) веществ.
Липофильные соединения проходят через БМ, растворяясь в ее липидах и двигаясь по
законам фиффузии в вязкой среде. Такой транспорт зависит от величины и формы
транспортируемого вещества (молекулы), а также, от вязкости БМ. Скорость
трансмембрального переноса в жирорастворимых веществах, имеет сильную
зависимость от температуры. Определив зависимость мембранной проницаемости для
тех или иных веществ от температуры, можно определить энергию активации их
транспорта через БМ. Экспериментально удалось установить, что величины энергии
активации для этиленгликоля составляют примерно 60 кДж/моль, для глицерина -
примерно 77 кДж/моль. Эти значения близко совпадают с энергией дегидратации
этих спиртов, что в свою очередь позволило предположить то, что прежде, чем
войти в липидный слой БМ, молекулы не электролитов подвергаются дегидратации.
Только после освобождения от водной оболочки они внедряются в
жидкокристаллическую структуру БМ и диффундируют в ней.
Молекулярные компоненты клеточной мембраны непрерывно перемещаются в
пределах своего слоя (латеральная миграция), задерживаясь на одном месте с. Поэтому существует мгновение,
когда свободное место оказывается еще незанятым другими компонентами
молекулярных мембран. Молекулы переносимого вещества могут занять это место под
действием градиентов. Разумеется, такое вторжение в жидкокристаллическую
структуру БМ возможно только в том случае, если размеры диффундирующей молекулы
укладываются в геометрические размеры вакантного узла (ячейки). Если же размеры
диффундирующих молекул соответствуют двум диаметрам ячейки, то, для продвижения
на один шаг, такая молекула должна выждать момент, когда одновременно станут
свободными две соседние ячейки, смежные с тем местом, где находится переносимая
молекула. Это событие также вероятно в БМ. Еще меньшую, но реальную вероятность
имеет одновременное образование смежных по соседству трех вакансий.
Следовательно, чем крупнее частицы (молекулы) проникающего через мембрану
вещества, тем меньше вероятность их проникновения сквозь БМ, при прочих равных
условиях. Это, прежде всего, отражается на скорости трансмембранного переноса
вещества. Так, крупные молекулы дольше ждут того момента, когда смогут
продвинуться на один шаг. Показателем скорости перемещения веществ через БМ,
может служить коэффициент диффузии данного вещества в БМ. Частицы диффундирующего
вещества перемещаются в БМ скачками от одного вакантного места к другому, при
этом, каждый раз выжидая образования вакантного узла. Такой механизм называется
прыжковым.
27. Транспорт
с участием переносчиков
Гидрофильные вещества практически не перемещаются в БМ за счет процессов
свободной диффузии. Транспорт многих гидрофильных веществ (моносахаридов,
аминокислот, некоторых ионов) обеспечивают подвижные переносчики. В частности,
пептид, имеющий циклическую структуру и находящийся в БМ, способен селективно
переносить ионы. Облегчение трансмембранного переноса обусловлено тем, что
вещество (например, глюкоза) преодолевает плазмолемму не путем свободной
диффузии через липидный бислой, а при помощи переносчика. В этой связи,
движение глюкозы сквозь мембрану эритроцита происходит быстрее, чем можно было
бы ожидать при ее свободной диффузии в мембранных липидах. В плазматической
мембране обнаружено несколько десятков транспортных белков, каждый из которых
переносит только определенное вещество. Переносчики в БМ могут работать,
используя различные виды перемещения:
миграционный;
ротационный;
сдвиговый.
Среди мигрирующих переносчиков можно выделить две разновидности. Одни
транспортеры мигрируют внутри мембран и взаимодействуют с переносимым веществом
только на ее поверхности; этот механизм транспорта называют малой
"каруселью". Другие мигрирующие переносчики способны покидать БМ и
выходить в примембранное пространство в поисках транспортируемого вещества.
Поиск направляется действием электростатических сил или химическим
взаимодействием. Вместе с переносимым веществом, транспортер второго типа
возвращается в БМ, проходит ее насквозь, выходит в противоположное
пространство, оставляя там переносимое вещество. Такой механизм называется
большой "каруселью". Тип "карусели" зависит от поверхности
активных свойств и растворимости самого переносчика. Как правило, по механизму
малой "карусели" работают транспортеры, плохо растворимые в воде и
являющиеся ПАВ. Схематически работу малой "карусели" можно
представить так:
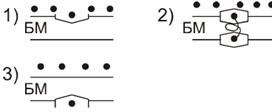
молекула
межклеточный информация связь
Миграционный механизм присущ переносчикам, размеры которых меньше толщины
БМ. Вместе с тем, транспортерами могут быть и более крупные белковые молекулы
или их комплексы, которые пронизывают БМ насквозь. Они переносят вещества через
БМ за счет поворота (ротации) или сдвига. Ротационный механизм заключается в
повороте крупной молекулы переносчика вокруг оси, лежащей в плоскости мембраны.
В результате чего, транспортируемые вещества (молекулы), посаженные на один
конец такого переносчика, оказываются на противоположной стороне БМ.
Ротационный механизм требует значительных затрат энергии и является эффективным
только в том случае, если одна молекула переносчика одновременно транспортирует
за один поворот много молекул переносимого вещества. Схематически работу
ротационного механизма можно представить в следующем виде:
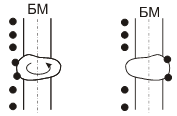
Более выгодным в энергетическом отношении является механохимический
процесс в молекуле переносчика, заключенный не в полном ее повороте, а в сдвиге
отдельных областей, относительно неподвижной части крупной молекулы. При этом,
вместе с участником переносчика, уходящим с поверхности вглубь мембраны,
передвигается транспортируемое вещество. Этот процесс напоминает движение лифта
в шахте. Графически это можно представить следующим образом:
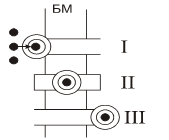
. Мембранные
каналы
Канал (пора), заполненный водой, насквозь пронизывает клеточную мембрану.
Длина канала, как правило, превосходит линейные размеры поперечного его
профиля, который имеет форму круга или неправильного эллипса. Конфигурация и
площадь поперечного сечения не одинаковы в различных местах одного и того же
канала. У большинства мембранных каналов максимальный линейный размер
поперечного профиля составляет 0,8 нм. Обычно, общая площадь каналов составляет
тысячную долю всей поверхности клеточной мембраны, но у некоторых клеток эта
площадь не превышает 0,000001 доли.
До настоящего времени неизвестны ни структура, ни состав липопротеиновых
комплексов, которые образуют мембранные каналы. Однако экспериментально
установлено, что скорость транспорта веществ по каналам определяется, главным
образом, числом каналов, приходящимся на единицу поверхности БМ. В этой связи,
проницаемость БМ при переносе веществ по каналам, рассчитывается по следующей
формуле:
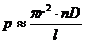
- усредненный радиус канала;
- число каналов на единицу поверхности БМ;
- коэффициент диффузии вещества в воде;
- длина канала, равная толщине БМ.
Скорость проникновения через плазмолемму нервных или мышечных волокон для
ионов 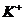 и
и 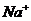 , и ряда других гидрофильных веществ,
примерно составляет
, и ряда других гидрофильных веществ,
примерно составляет 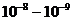 м/с, следовательно, их трансмембранный перенос осуществляется
на 5-6 порядков медленнее, чем свободная диффузия в водном растворе. Вероятно,
быстрее других проходят через каналы те гидрофильные вещества, диаметр которых
вместе с гидратной оболочкой близок к размеру поперечного профиля канала. Канал
представляет собой мембранный белок или белковый комплекс, полярные группы
которого направлены вовнутрь полости, являющийся характерным элементом -спирали. Полость канала заполнена
водой. Движение ионов по каналу строго однорядно, и его можно представить, как
последовательное замещение молекул воды из гидратной оболочки иона на полярные
группы, выстилающие внутреннюю поверхность канала. Ион, отдавая воду,
находящуюся в его гидратной оболочке, взаимодействует с полярными группами
канального белка и, подобно поршню, проталкивает молекулу воды, которая
встречается в канале на его пути.
м/с, следовательно, их трансмембранный перенос осуществляется
на 5-6 порядков медленнее, чем свободная диффузия в водном растворе. Вероятно,
быстрее других проходят через каналы те гидрофильные вещества, диаметр которых
вместе с гидратной оболочкой близок к размеру поперечного профиля канала. Канал
представляет собой мембранный белок или белковый комплекс, полярные группы
которого направлены вовнутрь полости, являющийся характерным элементом -спирали. Полость канала заполнена
водой. Движение ионов по каналу строго однорядно, и его можно представить, как
последовательное замещение молекул воды из гидратной оболочки иона на полярные
группы, выстилающие внутреннюю поверхность канала. Ион, отдавая воду,
находящуюся в его гидратной оболочке, взаимодействует с полярными группами
канального белка и, подобно поршню, проталкивает молекулу воды, которая
встречается в канале на его пути.
Мембранные каналы можно разделить на две группы:
потенциал зависимые (п-з);
потенциал независимые (п-нез).
Проницаемость каналов первой группы не просто зависит от трансмембранной
разности потенциалов, но и управляет ею. С функционированием потенциал
зависимых ионных каналов, связано важнейшее свойство живых тканей -
возбудимость. Таким образом, при пассивном транспорте, селективность БМ
определяется:
. Коэффициентом распределения веществ между липидами и водой.
. Избирательностью каналов или переносчиков по отношению к определенным
веществам.
. Биологические насосы. Активный транспорт
Активным транспортом называют транс-мембральный перенос веществ в
направлении, противоположном транспорту, который должен был бы происходить под
действием физико-химических градиентов. Активный транспорт направлен в сторону
более низкого электрохимического потенциала. Он необходим как для накопления в
клетках или органеллах веществ, в которых они нуждаются, даже из среды с их
низкой концентрацией, так и для выведения из клеток или органелл тех веществ,
содержание которых там должно поддерживаться на низком уровне, даже при
повышении этой концентрации в окружающей среде.
Из определения активного транспорта следует, что его важным свойством
является перенос веществ против действия физико-химических градиентов. Это
возможно только благодаря термодинамическому сопряжению биологических
процессов, причем, источником свободной энергии, необходимой для преодоления физико-химических
градиентов, могут служить продукты химического окисления. Их вклад в
термодинамическое сопряжение учитывается дополнительным слагаемым в
кинетических уровнях, которые описывают активный транспорт. Так, системы
уравнений для активного транспорта переноса ионов под действием
концентрационного и электронного градиентов:
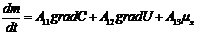
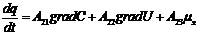
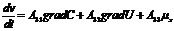
- число молей.
- (химический потенциал) количественно характеризует вклад
ферментативных реакций в свободную энергию БМ, которая необходима для
преодоления действия сопряжения градиентов (в данном случае, 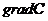 и
и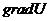 ). Вышеотмеченное позволяет
сформулировать второе характерное свойство активного транспорта - необходимость
энергетического обеспечения за счет свободной энергии, выделяющейся либо
непосредственно в ходе окислительно-восстановительных реакций, либо при
гидролизе АТФ, который синтезируется биологической системой впрок.
). Вышеотмеченное позволяет
сформулировать второе характерное свойство активного транспорта - необходимость
энергетического обеспечения за счет свободной энергии, выделяющейся либо
непосредственно в ходе окислительно-восстановительных реакций, либо при
гидролизе АТФ, который синтезируется биологической системой впрок.
Свободная энергия, затрачиваемая на трансмембранный перенос веществ
против концентрационного градиента, рассчитывается по формуле:
 , где
, где 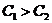
Так, чтобы откачать из клетки единичный заряд ионов в 1 Кл, необходимо затратить примерно
0,12 Дж.
У человека в состоянии покоя примерно 30-40 % всей энергии, образующейся
в ходе метаболических процессов, расходуется на активный транспорт. В некоторых
случаях, на его обеспечение может затрачиваться практически вся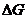 , которая выгребается клеткой. Так,
ткани, в которых активный транспорт особенно интенсивен, потребляют большее
количество кислорода, даже в состоянии покоя. Например, масса мозга человека
составляет примерно 1/50 массы тела, но, в условиях мышечного покоя, ткани
мозга поглощают примерно 1/5 часть всего кислорода, усвоенного организмом.
Общая мощность всех ионных насосов человеческого мозга составляет примерно 1
Вт.
, которая выгребается клеткой. Так,
ткани, в которых активный транспорт особенно интенсивен, потребляют большее
количество кислорода, даже в состоянии покоя. Например, масса мозга человека
составляет примерно 1/50 массы тела, но, в условиях мышечного покоя, ткани
мозга поглощают примерно 1/5 часть всего кислорода, усвоенного организмом.
Общая мощность всех ионных насосов человеческого мозга составляет примерно 1
Вт.
Третье свойство активного транспорта заключается в их специфичности.
Каждая из систем активного транспорта обеспечивает перенос через БМ только
данного вещества и не переносит другие. При этом, активный транспорт может быть
сопряжен с пассивным. Так, при активном транспорте ионов Na+ в том же
направлении переносятся, за счет пассивного транспорта, другие вещества
(глюкоза, аминокислоты). Это явление называют симпортом. Отдельная система
активного транспорта переносит одно вещество в одном направлении, а другое - в
противоположном. Так, (-) помпа закачивает ионы из межклеточной среды в цитоплазму и
откачивает ионы из клетки. Такой вид транспорта называют антипортом.
Необходимо отметить, что - насос не откачивает из клетки, а в нее. Когда эти ионы начинают
перемещаться через БМ по физико-химическим градиентам, при этом - помпа становится генератором АТФ.
Это явление получило название эффекта обращения активного транспорта. Так, на
перекачивание ионов против градиентов, насосы затрачивают свободную энергию,
гидрализуя АТФ, тогда, как при движении этих же ионов по градиентам, происходит
преобразование энергии градиентов в энергию макро энергетических связей
(синтезируется АТФ из АДФ).
Специфичность систем активного транспорта служит одним из самых
действующих механизмов селективной проницаемости клеточных мембран и придает им
векторные свойства.
30.
Компоненты систем активного транспорта
В любой системе активного транспорта веществ через биологические мембраны
можно выделить три основные компонента: источник свободной энергии, переносчик
данного вещества и сопрягающий (регулятор) фактор. Этот фактор сопрягает работу
переносчика с источником свободной. Все компоненты активного транспорта
образуют в БМ сложный молекулярный комплекс.
В большинстве систем активного транспорта, непосредственным источником
свободной энергии служит АТФ. За счет присоединения его концевой фосфатной
группы предварительно оторвавшейся при гидролизе, переносчик фосфолерируется и
преобретает за счет этого дополнительную энергию. Энергии достаточно для
преодоления физико-химических градиентов, которые препятствуют перемещению
веществ. Следовательно, фосфолерированный комплекс с переносимым веществом
способен преодолевать потенциальный барьер, который был непреодолимым до
фосфорелирования, и этот барьер препятствует переносу вещества через БМ.
Реже свободная энергия черпается непосредственно системами активного
транспорта из окислительно-восстановительных реакций, то есть, из цепи переноса
электронов. Эти системы называются редокспомпы. В качестве примера редокспомпы
можно рассмотреть перенос 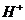 через внутреннюю мембрану митохондрий при клеточном дыхании.
через внутреннюю мембрану митохондрий при клеточном дыхании.
Вторым компонентом системы активного транспорта являются переносчики. В
разных системах активного транспорта переносчиками служат белковые молекулы,
которые работают по различным механизмам. Во-первых, переносчиком могут быть
сравнительно мелкие белковые молекулы, которые находятся в БМ. Эти переносчики
работают по механизму малой или большой карусели. Более крупные белковые
молекулы пронизывают БМ насквозь, и в этом случае, основным механизмом переноса
является ротационный или сдвиговый.
Третий компонент систем активного транспорта обеспечивает сопряжение
работы переносчика с источником свободной энергии. Такое сопряжение может
заключаться в процессах фосфорелирования АТФ, что имеет место при гидролизе.
Чтобы фосфорелировать переносчик, надо, прежде всего, гидролизовать АТФ.
Гидролиз АТФ достаточно эффективен только в присутствие специальных ферментов,
которые называются АТФазы. Они-то и служат факторами, которые сопрягают работу
переносчика и источника энергии. Для активного транспорта каждого вещества в
тех случаях, когда источником энергии является АТФ, обнаружена специфическая
АТФаза. Каждая из АТФаз активизируется тем веществом, чей транспорт она
обеспечивает. Например, 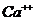 активируемая фаза, активируется только тогда, когда
концентрация
активируемая фаза, активируется только тогда, когда
концентрация 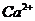 в примембранном пространстве достигает определенного уровня.
Превышение определенного уровня является сигналом к началу работы активного
транспорта.
в примембранном пространстве достигает определенного уровня.
Превышение определенного уровня является сигналом к началу работы активного
транспорта.
Все транспортные АТФазы связаны с клеточными мембранами и проявляют
высокую специфичность, катализируя реакции, течение которых зависит от
направления подхода в БМ транспортируемого вещества. Например, 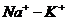 АТФаза преобразуется активно при
воздействии на нее внутри клетки, а - снаружи. Она не активизируется при
самых значительных концентрациях ионов Na в межклеточной среде (снаружи), а - в цитозоле (внутри). Зависимость
потока переносимого вещества через клеточную мембрану от его концентрации по
обе ее стороны при участии транспортной АТФазы, описывается следующим
выражением:
АТФаза преобразуется активно при
воздействии на нее внутри клетки, а - снаружи. Она не активизируется при
самых значительных концентрациях ионов Na в межклеточной среде (снаружи), а - в цитозоле (внутри). Зависимость
потока переносимого вещества через клеточную мембрану от его концентрации по
обе ее стороны при участии транспортной АТФазы, описывается следующим
выражением:
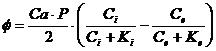
 - поток переносимого вещества;
- поток переносимого вещества;
 - концентрация АТФазы в биомембране;
- концентрация АТФазы в биомембране;
 - проницаемость БМ для комплекса фермент переносимого
вещества;
- проницаемость БМ для комплекса фермент переносимого
вещества;
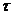 - концентрация переносимого вещества внутри клетки;
- концентрация переносимого вещества внутри клетки;
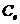 - концентрация переносимого вещества вне клетки;
- концентрация переносимого вещества вне клетки;
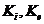 - коэффициенты диссоциативного комплекса внутри клетки и в межклеточной
среде соответственно.
- коэффициенты диссоциативного комплекса внутри клетки и в межклеточной
среде соответственно.
Таким образом, в клеточной мембране постоянно присутствует и переносчик и
АТФаза. В примембранном пространстве клетки находятся молекулы АТФ, выходящие
из митохондрий. Они подходят к местам активного транспорта. Однако, вся система
не работает до появления определенного стимула, которым обычно служит
наращивание концентрации вещества подлежащего к переносу. Этот фактор
активирует специфическую АТФазу, которая в свою очередь катализирует гидролиз
АТФ с отщеплением фосфатной концовой группы. Присоединенная к переносчику
фосфатная группа, фосфорилирует его. При фосфорилировании переносчик
приобретает дополнительную свободную энергию, необходимую и достаточную для
транспорта вещества через БМ, вопреки действиям физико-химических градиентов.
31. Системы
активного транспорта ионов
Система активного транспорта ионов (ионные насосы или ионная помпа)
обеспечивает неравновесное распределение ионов между клеткой и межклеточной
средой, а также различных органелл. Постоянство ионного состава цитозоля и
содержимого органелл, являются необходимым условием для поддержания
жизнедеятельности. Ионы входят в состав всех биологически важных молекул и
регулируют эффективность обмена веществ. Все превращения энергии регулируются
ионами. В организме они управляют сложным ионным процессом. Малейшее нарушение
баланса приводит к нарушению жизнедеятельности. Например, весь клеточный
метаболизм чрезвычайно чувствителен к изменению содержания в цитозоле. При его повышении
угнетается синтез белка и усиливается процесс образование липидов.
Следовательно, ионы Na выступают в роли регулировщика метаболизма клеточной
активности. Подобная функция характерна и для других ионов, в частности,
изменение содержания в цитозоли ионов водорода, , и др., сигнализирует ее
метаболическим системам о малейшем нарушении клеточной целостности, которая
достигается относительной обособленностью химического состава при наличии
многообразных связей со средой ее обитания.
В организме человека 50% содержится в межклеточной среде (интерстиции), 40% - в
костях, 10% - внутри клетки. В межклеточной среде катионам 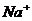 сопутствуют анионы
сопутствуют анионы 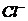 и бикарбоната, концентрация которых
значительно выше, чем в цитозоле. В отличие от ионов и
и бикарбоната, концентрация которых
значительно выше, чем в цитозоле. В отличие от ионов и 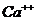 ионы
ионы  и
и 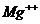 сосредоточены преимущественно внутри
клеток. Из 160 гр , входящего в состав тела человека среднего роста (170 см) и
среднего веса 70 кг, только 3 гр приходится на межклеточную среду. В цитозоле присутствует в ничтожно малой
концентрации, даже, в мышечных волокнах, где их содержание довольно велико, так
как там они сосредоточены не в цитозоле, а в "цистернах"
саркоплазматической сети, мембрана которых служит непреодолимым препятствием
для .
сосредоточены преимущественно внутри
клеток. Из 160 гр , входящего в состав тела человека среднего роста (170 см) и
среднего веса 70 кг, только 3 гр приходится на межклеточную среду. В цитозоле присутствует в ничтожно малой
концентрации, даже, в мышечных волокнах, где их содержание довольно велико, так
как там они сосредоточены не в цитозоле, а в "цистернах"
саркоплазматической сети, мембрана которых служит непреодолимым препятствием
для .
Стабильное поддержание ионного неравновесия, а также, перемещение ионов
через клеточную мембрану в сторону более низкого электрохимического потенциала,
для существования многих физиологических процессов, обеспечивается системами
активного транспорта.
32.
Калиево-натриевый насос
Благодаря системам активного транспорта для и , поддерживаются стабильные и высокие
градиенты концентрации этих ионов на плазмолемме любой клетки. Разница в их
молекулярной концентрации между цитозолем и межклеточной средой достигается
10-20 раз. При этом, больше в цитозоли, - в межклеточной среде. При действии
концентрационных градиентов входят в нее. Пассивный транспорт привел бы к ликвидации
ионного неравновесия, но этому препятствует работа 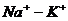 насоса. Он откачивает из цитозоля в межклеточную среду и
закачивает в клетку. насос - энергоемкая система.
насоса. Он откачивает из цитозоля в межклеточную среду и
закачивает в клетку. насос - энергоемкая система.
Компонент помпы является АТФ (источник энергии) АТФазы, которые одновременно служит и
переносчиком и сопрягающим фактором. Молекулярный механизм использования
энергии АТФ для работы помпы еще полностью не понят, однако, предполагается, что на
внутренней стороне БМ находятся молекулярные комплексы, способные
фосфорелировать за счет присоединения фосфатной группы АТФ. Фосфатная группа
отщепляется от АТФ при гидролизе. Фосфорелирующий транспортный комплекс
переносит, связанный с ним на наружную сторону клеточной мембраны, где обменивает на . комплекс транспортирует его внутрь
клетки, где он фосфолерируется, то есть, отщепляется фосфатная группа, так как
вся свободная энергия истрачена. Для следующего транспортного цикла комплексу
необходимо новое фосфорелирование за счет гидролиза АТФ.
В настоящее время есть несколько гипотез, конкретизирующих участие в
работе комплекса -АТФазы. Однако, ни одна из них не является общепринятой, но,
все модели активного транспорта и предусматривает конфирмацию перестройки -АТФазы. Эти перестройки
осуществляются в пространстве БМ.
Некоторые условно полагают, что, будто помпа действует по принципу
перистальтического насоса, который переменно открывает и закрывает и , расположенные в БМ по соседству с АТФазой.
Ее конформация перестройки обуславливается чередованием фосфорелирования
и дефосфорелирования, обеспечивают изменение проницаемости ионов каналов, при
этом, эти насосы работают в противофазе, то есть, когда канал открыт, - закрыт, и наоборот. Согласно этой
гипотезе, АТФаз служит не как переносчик, а своеобразное планирование
ионных каналов.
Другая гипотетическая схема действия помпы предполагает, что транспорт
АТФаз работает, как переносчик. Молекула этого фермента пронизывает плазмолемму
насквозь, контактируя одним полюсом с цитоплазмой, другим - с межклеточной
средой. В при мембранных участках цитоплазмы находятся митохондрии, из которых
выходят молекулы АТФ. АТФ выступает в контакте с БМ, следовательно, и с АТФазой. Содержание ионов Na в
цитоплазме поддерживается на строго определенном уровне. АТФаза активируется ионами и , но проявляется при этом ярко
выраженная векторность. Ионы действуют на нее только со стороны цитоплазмы, а - только из межклеточной среды.
Вместе с тем, ее специфичность по отношению к выше, чем к , так, для включения в действие АТФазы, незаменимы, в то время, как можно заменить одновалентным
катионом, причем, с рубидием транспортная система функционирует лучше, чем с . По-видимому, для ферм. характерны
различные механизмы распознавания на цитоплазме стороне БМ и других
катионов на ее наружной поверхности.
Экспериментально установлено, что за счет гидролиза одной молекулы АТФ
осуществляется активный транспорт трех ионов и двух ионов , то есть, их сопряженному трансмембранному
переносу свойственна стехиометрия (3/2). Она сохраняется вне зависимости от
величины и направления концентрационного и электрического градиентов.
Стехиометрия обусловливает электрогенные свойства помпы. Перенос двух внутрь клетки и трех ионов Na из
нее, создает разность потенциалов, относительно межклеточной среды. При этом
цитозоль имеет отрицательный заряд относительно межклеточной среды (внутри
клетки анионов больше, чем катионов).
насос угнетается различными агентами, из которых наиболее
активны следующие:
. Сердечные гликозиды.
. Избыток ионов внутри клетки.
. Дыхательные яды.
Дыхательные яды блокируют окислительное фосфорилирование в митохондриях
и, соответственно, они останавливают работу любой системы активного транспорта,
то есть, в их действии нет специфичности, так как они лишают свободной энергии
активного транспорта. Специфичным плантатором насоса служит строфангин Г (уабаин),
который является ингибитором АТФазы. Даже в концентрации, равной м/а, уабаин подавляет активность помпы на 50%. Под действием
гликозидов подавляется выведение из клеток, и приток в них, тогда как пассивный выход из цитозоли не изменяется.
. Кальциевый насос (КН)
КН обеспечивает стабильно низкий уровень 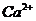 в цитозолях, в отличие от насоса, он выводит избыток ионов , не в межклеточную среду, а в
органеллы (главным образом, в эндоплазматическую сеть). Поэтому, основные места
локализации насоса находятся в большинстве типов клеток на внутренней мембране,
а не на плазмолемме (что имело место для насоса). Достаточно детально изучена
работа помпы в мембранах саркоплазматической
сети, где ее активность наиболее высока.
в цитозолях, в отличие от насоса, он выводит избыток ионов , не в межклеточную среду, а в
органеллы (главным образом, в эндоплазматическую сеть). Поэтому, основные места
локализации насоса находятся в большинстве типов клеток на внутренней мембране,
а не на плазмолемме (что имело место для насоса). Достаточно детально изучена
работа помпы в мембранах саркоплазматической
сети, где ее активность наиболее высока.
Источником энергии для системы активного транспорта ионов служит АТФ. Вторым и третьим
компонентом работы насоса является активированная АТФаза. Кроме того,
для работы АТФазы необходимо присутствие ионов 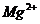 . В саркоплазматической цепи на долю АТФазы приходится около 60% общего
мембранного белка. Предполагается, что в мембране саркоплазматической сети нет
другого белка, кроме АТФазы. Остальные 40% белков принадлежат периферическим
протеинам. На перенос двух ионов затрачивается одна молекула АТФ, но, при
очень высоких физико-химических градиентах, это отношение изменяется с двух к
одному, до одного к одному.
. В саркоплазматической цепи на долю АТФазы приходится около 60% общего
мембранного белка. Предполагается, что в мембране саркоплазматической сети нет
другого белка, кроме АТФазы. Остальные 40% белков принадлежат периферическим
протеинам. На перенос двух ионов затрачивается одна молекула АТФ, но, при
очень высоких физико-химических градиентах, это отношение изменяется с двух к
одному, до одного к одному.
В механизме действия насоса выделяется три этапа:
Во-первых, сигналом к активному транспорту служит увеличение допустимого
уровня ионов в цитозоле. Установлено, что постоянная связывания ионов
кальция - АТФазой, имеет порядок л/моль, то есть, адсорбция ионов на транспортном ферменте происходит
уже при концентрации ионов в цитозоли примерно моль/л. - АТФаза связывает не только ионы, но и ионы . Центры захвата ионов и АТФ локализованы на поверхности
фермента. Эта поверхность обращена к цитозоле.
Активированная ионами транспортная АТФаза катализирует гидролиз АТФ, что является
основным событием второго этапа в работе помпы. При гидролизе АТФ, от нее
отщепляется концевая фосфатная группа и присоединяется к - АТФазе, которая приобретает при
этом дополнительную свободную энергию равную примерно 37,4 Дж/моль. За счет
этой энергии образуется фермент-фосфатный комплекс.
Третий этап работы насоса - переход ионов на противоположную сторону внутри
БМ, что связано с изменением конформации АТФазы. Изменение конформации
приводит к перемещению молекулы фермента в пространстве БМ. Предполагается, что
происходит сдвиг подвижных групп АТФазы, на которой адсорбированы
ионы. На противоположной стороне БМ
транспортная АТФаза освобождается от ионов, так как ферментно-фосфатный
комплекс гидрализуется, после того, как затрачивает полученную энергию на
активный транспорт ионов. Вслед за гидролизом ферментно-фосфатного комплекса,
происходит дефосфорилирования фермента. Возвращение - связывающих центров в исходное
состояние является следствием восстановительной конформации молекулы АТФазы. Это состояние переходит в то,
которое ей является свойственным в нефосфорелированном состоянии.
насос, в отличие от насоса, не проявляет электрогенных
свойств, то есть, активный транспорт ионов не сопровождается образованием
разности потенциалов на БМ. Неэлектрогенность помпы обусловлена высокой
проницаемостью этой мембраны для многих ионов. В этой связи, мембранный
потенциал, который создается переносом ионов, сразу же падает из-за утечки других
ионов.
34. Механизмы
биоэлектрогенеза и его роль в возбуждении. Физико-химические свойства
биоэлектрогенеза (БЭГ)
Живая ткань обладает не только пассивными, но и активными электрическими
свойствами, являясь источниками электромагнитной энергии. Природа БЭГ была
установлена только после появления теории электрической диссоциации, которая
была разработана в 1887 г. Аррениусом. Живые ткани обладают свойствами
электролита, которые диссоциируются на катионы и анионы. В этой связи, самой
простой моделью источника электромагнитной энергии биологических тканей служит
концентрационный элемент Нернста. В нем растворы соли разной концентрации
разделены мембраной, имеющей неодинаковую проницаемость для катионов и анионов,
на которые диссоциирует данная соль. Вследствие существования разности
концентрации, электролит стремится диффундировать через мембрану. Но, в силу
неодинаковой проникающей способности образующих его ионов, один из них
преодолевает мембрану, а другой задерживается ею. В результате, на мембране
образуется двойной слой зарядов. При этом, более разбавленный раствор принимает
заряд того иона, который лучше проникает сквозь мембрану. Если это катион, то
электрический ток во внешней цепи концентрационного электролита течет от
электрода, погруженного в раствор с меньшей концентрацией, к электроду,
находящемуся в более крепком растворе. При одинаковой проникающей способности
катионов и анионов, на которые диссоциирует электролит, электрическая энергия
не генерируется, как бы ни был высок концентрационный градиент на мембране.
Схематически электролит Нернста можно представить:
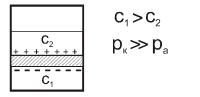
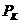 - проницаемость катионов
- проницаемость катионов
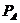 - проницаемость анионов
- проницаемость анионов
Электродвижущая сила, возникающая в концентрированном электролите,
который образован раствором одной соли, определяется из уравнения Нернста:
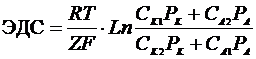
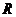 - универсальная газовая постоянная;
- универсальная газовая постоянная;
- температура;
- валентность ионов;
- число Фарадея;
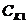 - концентрация анионов в первой области;
- концентрация анионов в первой области;
- проницаемость для катионов через мембрану;
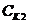 - концентрация катионов во второй области;
- концентрация катионов во второй области;
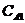 - концентрация анионов в первой области;
- концентрация анионов в первой области;
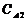 - концентрация анионов во второй области;
- концентрация анионов во второй области;
 - проницаемость для анионов через мембрану.
- проницаемость для анионов через мембрану.
Уравнение Нернста явилось результатом математической обработки многочисленных
результатов экспериментального исследования функциональной зависимости ЭДС
концентрационного элемента от соотношения концентрации соли в растворах,
разделяемых мембраной. По Нернсту, десятикратная разность концентраций
однозарядного иона создает ЭДС примерно 60 мВ (а двухзарядные ионы создают ЭДС
примерно 30 мВ), следовательно, одновалентные ионы вносят наибольший вклад в
БЭГ.
Наличие концентрационных градиентов является необходимым, но
недостаточным условием БЭГ. Он обусловливается также тем, что сквозь БМ катионы
проникают лучше, чем анионы. Концентрационные градиенты стремятся выровнять
содержание всех ионов по обе стороны БМ. Однако, БМ препятствует этому
процессу, поддерживая тем самым ионную асимметрию. Важный вклад в поддержание
ионной асимметрии и соответствующих концентрационных градиентов (например,
ионов и ) на плазмолемме любой клетки вносит
их активный транспорт.
Таким образом, при наличии БЭГ всегда соблюдается два обязательных
условия:
. Существование концентрационных градиентов электролитов на клеточной
мембране.
. Наличие неодинаковой проницаемости этой мембраны для катионов и
анионов, на которые диссоциируют электролиты в живой клетке (внутри и вне
клетки).
При рассмотрении живых тканей в качестве концентрационного элемента,
необходимо учитывать диффузию через БМ не одного иона, а всех, концентрации
которых неодинаковы внутри и вне клетки, и которые способны при этом проникать
через БМ.
35. Потенциал покоя
Уравнение Гольдмана позволяет рассчитывать разность потенциалов,
существующую у разных клеток между цитоплазмой и межклеточной средой в
состоянии покоя и в состоянии возбуждения. Это уравнение имеет вид:
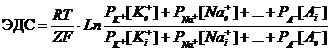
ЭДС в состоянии покоя называют потенциалом покоя (ПП),
в состоянии возбуждения - потенциалом действия (ПД). Результаты расчетов
довольно близко совпадают с результатами экспериментов. Наилучшее совпадение
имеет место для ПП.
Анализ результатов эксперимента показал, что в состоянии покоя
проницаемость для больше проницаемости для  и
и 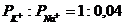 . Это позволяет считать проницаемости
для ионов и анионов бесконечно малыми величинами по отношению к
проницаемости для . В этом случае численное значение потенциала покоя можно
рассчитывать по формуле:
. Это позволяет считать проницаемости
для ионов и анионов бесконечно малыми величинами по отношению к
проницаемости для . В этом случае численное значение потенциала покоя можно
рассчитывать по формуле:
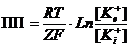
Необходимо отметить, что ПП возник не за счет более быстрой диффузии
ионов по сравнению с . Разница между проницаемостями и определяет только направление
катионного потока, который создает ПП на БМ. Поскольку содержание в цитоплазме больше, чем в
межклеточной среде, то катионный поток направлен из клетки наружу (выходящий
ток). Соотношение концентрации ионов в межклеточной среде и внутри клетки
определяет амплитуду ПП. Причиной возникновения трансмембранной разности
потенциалов в состоянии покоя, является скорость диффузии через БМ ионов и анионов высокомолекулярных
органических веществ, находящихся в цитозоли. Катионы проходят сквозь плазмолемму, а анионы
задерживаются ею, что приводит к образованию не плазмоалемме двойного
электрического слоя зарядов. При этом, межклеточная среда, как более
разбавленный раствор, приобретает "+" заряд, а цитоплазма
"-".
Расчеты экспериментальных данных свидетельствуют о том, что все клетки
организма в состоянии покоя характеризуются определяемым потенциалом
(поляризацией). Клеточная мембрана всегда заряжена и всегда на ее внутренней
поверхности поддерживается "-" заряд (разность потенциала), а на
внешней - "+" заряд. Трансмембранная разность потенциалов в разных
клетках различна, но всегда составляет несколько десятков мВ. Так, например,
для гигантского аксона кальмара потенциал покоя составляет примерно -85 мВ, в
то время, как для мембран эпителиальных клеток ПП - в 2-3 раза ниже.
Незначительная на первый взгляд величина потенциала покоя создает высокий
градиент потенциала на плазмолемме, так как толщина БМ составляет примерно 10
нм, следовательно, напряженность электрического поля на плазмолемме может
достигать В/м.
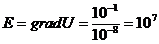
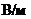
Избирательная проницаемость плазмолеммы каждой клетки обеспечивает
стабильное разобщение катионов и анионов, вследствие чего, на ней стойко
поддерживается высокий градиент потенциала, и она обладает ЭДС. Эта ЭДС направлено
против причины ее вызывающей и противодействует дальнейшему разобщению зарядов
на БМ. Существование ионных градиентов на БМ связано с работой системы
активного транспорта и является энергоемким процессом. В этой связи, потенциал
покоя частично экономит энергетические затраты свободной энергии на поддержание
этих градиентов, так как потенциал покоя действует в противоположном
направлении концентрационному градиенту.
. Потенциал действия
При действии на живые ткани раздражителей, изменяются условия генерации
ЭДС в клеточных концентрационных элементах, что выражается в возникновении ПД.
Согласно натриевой теории (гипотезе), при возбуждении нервных или мышечных
волокон, изменяется проницаемость плазмолеммы клеточных структур для ионов и . Мгновенно, на короткий промежуток
времени, проницаемость для ионов и становится равным, как 1:30.

При этом, проницаемость для оказывается намного больше
проницаемости для анионов, с которыми ион образует соли в межклеточной среде.
Следовательно, в момент возбуждения, проницаемость для ионов намного больше проницаемости для
ионов и анионов. Это позволяет принять проницаемости
для ионов 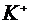 и анионов за бесконечно малые величины. После преобразования
уравнения Гольдмана, получаем формулу для расчета так называемого натриевого
потенциала. Он равен превышению по абсолютной величине потенциала действия над
потенциалом покоя.
и анионов за бесконечно малые величины. После преобразования
уравнения Гольдмана, получаем формулу для расчета так называемого натриевого
потенциала. Он равен превышению по абсолютной величине потенциала действия над
потенциалом покоя.
Разность модулей ПД и ПП называют потенциалом инверсии. В аксоне он
эквивалентен натриевому потенциалу, то есть:

Численное значение потенциала инверсии неодинаково в разных клетках, но
всегда составляет десятки мВ, и знак этого потенциала положительный,
относительно потенциала покоя:
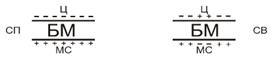
Как правило, потенциал 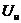 меньше ПП по модулю. Так, например, в гигантском аксоне
кальмара примерно равен +55 мВ, а в
миакардиальных волокнах он составляет примерно +20 мВ (для сердца человека). ПД
определяется суммой абсолютных значений ПП и .
меньше ПП по модулю. Так, например, в гигантском аксоне
кальмара примерно равен +55 мВ, а в
миакардиальных волокнах он составляет примерно +20 мВ (для сердца человека). ПД
определяется суммой абсолютных значений ПП и .
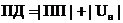
Продолжительность существования ПД в большинстве нервных и мышечных
волокон невелика (в пределах долей единиц мс.), причем, не требуется никаких
воздействий, чтобы нейрон или мышечное волокно вернулось к исходному состоянию
поляризации, то есть, приобрело обычный ПП. Возбудимый участок цитоплазмы
приобретает положительный потенциал, относительно межклеточной среды, и такое
состояние называют деполяризацией. Возвращение к прежней поляризации называют
реполяризацией.
ПД создается за счет более быстрой диффузии сквозь плазмолемму ионов , по сравнению с анионами, которые
образуют с анионами соли в межклеточной среде, следовательно, деполяризация
(появление ) связана с вхождением в цитоплазму ионов .
Возникновение ПД обусловлено не нарушением ионных концентраций в
цитозоли, а падением электрического сопротивления плазматической мембраны, вследствие
повышения ее проницаемости для ионов . Так, при деполяризации
сопротивление плазмолеммы уменьшается примерно в 40 раз. Входящий поток ионов является результатом его пассивного
транспорта по мембранным каналам под действием концентрационного и
электрического градиентов, а выходящий поток ионов обеспечивается работой натриевой
помпы (активный транспорт). В состоянии покоя, встречные потоки ионов уравновешенны, в то время, как при
возбуждении в течение времени существования ПД, система встречных потоков
нарушается, и ионы устремляются из межклеточной среды внутрь клетки. При этом
система активного транспорта оказывается неспособна моментально ликвидировать
(компенсировать) резкое усиление входящего потока. Она делает это с некоторым
запозданием, в фазу реполяризации. Мембранные потенциалы являются не
равновесными, а стационарными, так как они поддерживаются в условиях
существования встречных потоков через БМ. Сдвиги мембранных потенциалов связаны
с нарушением действия стационарного режима, причем, возбуждение сопровождается
усилением входящего и выходящего потоков ионов . Следовательно, возбуждение не
включает систему активного транспорта ионов , а напротив, активизирует ее,
однако, даже при максимальной активизации работы натриевой помпы, она не может
воспрепятствовать кратковременному накоплению небольшого количества ионов в цитоплазме. В промежутках между
последовательными генерациями ПД (в состоянии покоя)  помпа откачивает из цитоплазмы
избыток ионов , который образовался при возбуждении, и ликвидирует
созданный дефицит ионов .
помпа откачивает из цитоплазмы
избыток ионов , который образовался при возбуждении, и ликвидирует
созданный дефицит ионов .
37.
Возбудимые и невозбудимые мембраны
Все клеточные БМ можно разделить на возбудимые (электрогенные) и
невозбудимые (неэлектрогенные). Для неэлектрогенных БМ присущ только ПП, а для
возбудимых БМ присущи как ПП, так и ПД. Указанное различие обусловлено
присутствием в БМ потенциал зависимых ионных каналов. В невозбудимых
биологических мембранах находятся только потенциал независимые ионные каналы, а
в возбудимых присутствуют и те, и другие. В физиологии свойства возбудимости не
ограничивают мембранами, а распространяют на ткани, подразделяя их на
возбудимые и невозбудимые. Считают возбудимыми такие ткани, клеточная мембрана
которых обладает потенциал зависимыми ионными каналами. К ним относят нервную и
мышечную ткани. Только в них под действием раздражителей возникают ПД, причем,
они сопровождаются целым комплексом других процессов, также, развивающихся в
ответ на стимуляцию (на раздражение). Среди них можно отметить: изменение
обмена веществ, изменение температуры, изменение электрического импеданса, а
также, специфические реакции, например, проведение нервных импульсов и
сокращение мышечных волокон.
Способность возбудительной ткани отвечать таким образом на действия
раздражителей, называют возбудимостью - . Ее количественной мерой служит
интенсивность порогового раздражителя 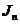 , то есть, самого слабого стимула, в
ответ на который возникает ПД, а вслед за ним и специфическая реакция. Чем ниже
интенсивность , тем выше .
, то есть, самого слабого стимула, в
ответ на который возникает ПД, а вслед за ним и специфическая реакция. Чем ниже
интенсивность , тем выше .
Следовательно, между ними существует обратно пропорциональная
зависимость:
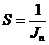
Реакцию возбудимой ткани на раздражение называют возбуждением. У
возбуждения много различных проявлений, но одним из важнейших является ПД,
поскольку он проявляется при любых специфических реакциях. Плазматические
мембраны нейрона или мышечного волокна возбудимы не на всем протяжении. На
поверхности этих микроструктур, участки возбудимых и невозбудимых БМ
чередуются, так как не везде в плазмолемму встроен потенциал зависимый ионный
канал.
Существенные различия между возбудимыми и невозбудимыми БМ проявляются и
из ВАХ. Для невозбудимых БМ ВАХ имеет вид:

Из ВАХ видно,
что при изменении мембранного потенциала от уровня ПП (-85 мВ) до 0, ее
электропроводность остается неизменной. С понижением абсолютного значения
мембранного потенциала, плотность тока сквозь невозбудимые БМ пропорционально
уменьшается.
ВАХ возбудимых БМ не линейна, и имеет следующий вид:

При понижении трансмембранной разности потенциалов до определенного
значения, плотность тока падает до 0, а затем, дальнейшее уменьшение
абсолютного значения мембранного потенциала приводит к резкому нарастанию
плотности тока, который к тому же изменяет свое направление (с выходящего из
волокна на входящее в него). Уровень мембранного потенциала, который
соответствует излому на ВАХ, называют критическим мембранным потенциалом (КМП).
На рисунке КМП равен -55 мВ, что характерно для гигантского аксона кальмара.
ВАХ свидетельствует о том, что деполяризация возбудимой мембраны, начиная с
уровня КМП, вызывает изменение мембраной электропроводности.
ВАХ возбудимой БМ имеет N-образную форму, при этом, особое внимание
заслуживает второй участок ВАХ. Его называют участком отрицательного
диффузионного сопротивления (ОДС). Поскольку, при падении абсолютного значения
U, I может нарастать только в том случае, если изменение напряжения
сопровождается понижением сопротивления. Такая аномалия ВАХ возбудимой мембраны
обусловлена присутствием в ней потенциал зависимых ионных каналов. Дело в том,
что плотность трансмембранного ионного тока может определяться, в принципе,
только двумя факторами:
. Числом открытых каналов.
. Электропроводностью каждого из них.
Известно, что электропроводность отдельного канала равна либо 0, при
закрытых воротах, либо максимальному значению, порядка 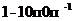 , что соответствует переносу
, что соответствует переносу 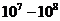 ионов за 1 секунду, при открытых
воротах. Не бывает промежуточных значений электропроводности отдельного канала,
поэтому, изменение сопротивления возбудимой мембраны зависит от количества
открытых каналов, при данном уровне мембранного потенциала. Сенсоры напряжения
различных потенциал зависимых ионных каналов, даже, если они принадлежат одному
типу, например, иону , обладают неодинаковой чувствительностью к сдвигам
трансмембранной разности потенциалов, то есть, у них неодинаков порог
срабатывания, поэтому, в разных каналах воротные процессы включаются при
различном уровне мембранного потенциала. Очевидно, что КМП соответствует такой
деполяризации, при которой начинают открываться наиболее чувствительные
потенциал зависимые натриевые каналы данной БМ. Начавшееся по ним движение
ионов усиливает деполяризацию, которая, по
мере нарастания, открывает все новые и новые потенциал зависимые натриевые
каналы до тех пор, пока все они не перейдут в открытое состояние.
ионов за 1 секунду, при открытых
воротах. Не бывает промежуточных значений электропроводности отдельного канала,
поэтому, изменение сопротивления возбудимой мембраны зависит от количества
открытых каналов, при данном уровне мембранного потенциала. Сенсоры напряжения
различных потенциал зависимых ионных каналов, даже, если они принадлежат одному
типу, например, иону , обладают неодинаковой чувствительностью к сдвигам
трансмембранной разности потенциалов, то есть, у них неодинаков порог
срабатывания, поэтому, в разных каналах воротные процессы включаются при
различном уровне мембранного потенциала. Очевидно, что КМП соответствует такой
деполяризации, при которой начинают открываться наиболее чувствительные
потенциал зависимые натриевые каналы данной БМ. Начавшееся по ним движение
ионов усиливает деполяризацию, которая, по
мере нарастания, открывает все новые и новые потенциал зависимые натриевые
каналы до тех пор, пока все они не перейдут в открытое состояние.
Следовательно, при изменении напряжения на аксолемме от -55 до -10 мВ (в
нашем случае), потенциал зависимые натриевые каналы работают, как системы с
положительной обратной связью по току. Стоит запустить в действие наиболее
чувствительные компоненты такой системы, чтобы в дальнейшем вся она вовлеклась
в электрогенез. Это свойство любой возбудимой мембраны. Однако, конкретные
значения мембранных потенциалов, соответствующие началу и концу действия,
"+" обратной связи в разных мембранах различны.
После того, как все каналы откроются, чему соответствует максимальное
значение тока ионов ,через возбудимые мембранные ворота натриевых каналов
начинают закрываться, и ток падает до определенного значения.
38.
Рефракторность
Процесс возбуждения сопровождается изменением возбудимости БМ.
Рефрактерность - это слово, в переводе означающее
"невпечатлительность". Рефрактерность - это изменение возбудимости
при возбуждении. Динамику возбудимости при возбуждении во времени, можно
представить в следующем виде:

АРФ - абсолютная рефракторная фаза;
ОРФ - относительная рефракторная фаза;
ФЭ - фаза экзальтации.
На кривой выделено три участка, которые называют фазами.
Развитие возбуждения в начале сопровождается полной утратой возбудимости
(S=0). Это состояние называют абсолютной рефрактерной фазой (АРФ). Оно
соответствует времени деполяризации возбудимой мембраны, то есть, переход
мембранного потенциала от уровня ПП до пикового значения ПД (до максимального
значения 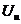 ) (см. ПД). В течение АРФ, возбудимая
мембрана не может генерировать новый ПД, даже, если на нее подействовать сколь
угодно сильным раздражителем. Природа АРФ состоит в том, что во время
деполяризации все потенциал- зависимые ионные каналы находятся в открытом
состоянии, и дополнительные стимулы (раздражители) не могут вызвать воротные
процессы, так как им просто не на что действовать.
) (см. ПД). В течение АРФ, возбудимая
мембрана не может генерировать новый ПД, даже, если на нее подействовать сколь
угодно сильным раздражителем. Природа АРФ состоит в том, что во время
деполяризации все потенциал- зависимые ионные каналы находятся в открытом
состоянии, и дополнительные стимулы (раздражители) не могут вызвать воротные
процессы, так как им просто не на что действовать.
АРФ изменяется относительной рефрактерной фазой (ОРФ), в течение которой,
возбудимость от 0 возвращается к исходному уровню (S=So). ОРФ совпадает по
времени с реполяризацией возбудимой мембраны. В течение этого времени, все
большее число потенциалзависимых каналов завершает воротные процессы, с
которыми было связано предыдущее возбуждение. При этом каналы вновь обретают
способность к следующему переходу из закрытого состояния в открытое, под
действием очередного стимула. Во время ОРФ пороги возбуждения постепенно
снижаются и, следовательно, возбудимость восстанавливается до исходного уровня
(до Sо).
За ОРФ следует фаза экзальтации (ФЭ), для которой характерно повышенная
возбудимость (S>So). Она, очевидно, связана с изменениями свойств сенсора
напряжения во время возбуждения. Предполагается, что за счет конформационных
перестроек белковых молекул изменяется их дипольный момент, что приводит к
повышению чувствительности сенсора напряжения и к сдвигам мембранной разности
потенциалов, то есть, критический мембранный потенциал как бы приближается к
ПП.
Разные мембраны имеют неодинаковые продолжительности каждой фазы. Так,
например, в скелетных мышцах АРФ в среднем длится 2,5 мс, ОРФ - порядка 12 мс,
ФЭ - 2 мс. Миокард человека отличается очень долгой АРФ, равной 250-300 мс, что
обеспечивает четкую ритмичность сердечных сокращений. Различие во времени
каждой фазы объясняется тем, какие каналы ответственны за этот процесс. В тех
мембранах, где возбуждаемость обеспечивается натриевыми каналами, рефрактерные
фазы наиболее быстротечны, и ПД имеет наименьшую продолжительность. Если же, за
возбуждаемость ответственны кальциевые каналы, то рефрактерные фазы
затягиваются до секунд. В мембране миокарда человека присутствуют и те, и
другие каналы ( и 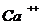 ), вследствие чего, длительность рефракторных фаз занимает
промежуточное значение.
), вследствие чего, длительность рефракторных фаз занимает
промежуточное значение.
39. Распространение возбуждения
Возбудимая мембрана относится к нелинейным и активным средам. Активной
называют такую среду, которая генерирует электромагнитную энергию под действием
приложенного к ней электромагнитного поля. Способность к БЭГ (к образованию ПД)
отображает активный характер возбудимости мембраны. Активный характер
проявляется также и в наличии участка ОДС на ее ВАХ. Это же свидетельствует о
нелинейности возбудимой мембраны, поскольку, отличительным признаком
нелинейности среды служит нелинейная функция, зависимость потоков от сил,
которые их вызывают. В нашем случае - это зависимость ионного тока от трансмембранного
напряжения. Применительно к электрическому процессу в целом, это означает
нелинейную зависимость тока от напряжения.
Нервные и мышечные волокна, будучи генераторами ЭМЭ (электромагнитной
энергии), обладают и пассивными электрическими свойствами. Пассивные
электрические свойства характеризуют способность живых тканей поглощать энергию
внешнего ЭМП (электромагнитного поля). Эта энергия затрачивается на их
поляризацию, и она характеризуется потерями в тканях. Потери в живых тканях
приводят к затуханию ЭМП, то есть, говорят о декременте. Закономерности
затухания ЭМП идентичны для потенциалов, приложенных извне, и генерируемых
самими живыми тканями (ПД). Степень декремента (затухания) зависит от
сопротивления и емкости ткани. В электронике, сопротивление и емкость
(индуктивность) называют пассивными свойствами электрических цепей.
Допустим, что в какой-то точке БМ потенциал мгновенно возрос до величины , в результате, затухания потенциал
будет уменьшаться по exp закону:
, в результате, затухания потенциал
будет уменьшаться по exp закону:
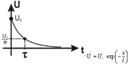
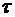 - постоянная времени затухания, то есть, время, в течение
которого амплитуда уменьшается в e раз (37%).
- постоянная времени затухания, то есть, время, в течение
которого амплитуда уменьшается в e раз (37%).
Постоянная времени зависит от пассивных свойств нервных или мышечных волокон:

Так, например, для гигантского аксона кальмара, Rн составляет примерно  , а
, а 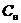 равно примерно
равно примерно , следовательно, равна примерно 1 мс.
, следовательно, равна примерно 1 мс.
Угасание потенциала происходит не только с течением времени в точке его
возникновения, но и также, при распределения потенциала вдоль БМ, по мере
удаления от этой точки. Такой декремент является функцией не времени, а
расстояния:
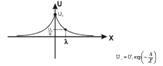
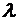 - постоянная длины, то есть, это расстояние, на которое
- постоянная длины, то есть, это расстояние, на которое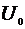 уменьшается в
уменьшается в  раз.
раз.
Декремент потенциала вдоль БМ происходит достаточно быстро в обе стороны
от того места, где возник скачок мембранного потенциала. Распределение
электрического потенциала на БМ устанавливается практически мгновенно, так как
скорость распределения ЭМП близка к скорости распространения света (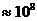 м/с). С течением времени, потенциал
падает во всех точках волокна (мышечного или нервного). Для длительно
существующих сдвигов мембранного потенциала, постоянная длины вычисляется по
формуле:
м/с). С течением времени, потенциал
падает во всех точках волокна (мышечного или нервного). Для длительно
существующих сдвигов мембранного потенциала, постоянная длины вычисляется по
формуле:
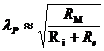
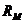 - погонное сопротивление мембраны (
- погонное сопротивление мембраны (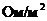 );
);
 - сопротивление цитоплазмы (Ом);
- сопротивление цитоплазмы (Ом);
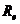 - сопротивление межклеточной среды (Ом).
- сопротивление межклеточной среды (Ом).
При коротких импульсах, как ПД, необходимо учитывать емкостные свойства
БМ. Из экспериментов установлено, что емкость БМ вносит искажение в эту
формулу. С учетом поправки, постоянная длины для ПД, оценивается величиной.
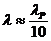
Чем больше , тем слабее декремент потенциала вдоль мембраны. Так, в
гигантском аксоне кальмара примерно равна 2,5 мм. У больших волокон составляет примерно 10-40 их
диаметров.
Таким образом, и являются основными параметрами, которые характеризуют
кабельные свойства БМ. Они количественно определяют декремент потенциала, как
во времени, так и в пространстве. Для уяснения механизмов распределения
возбуждения, особо важное значение имеет волокон. Анализ кабельных свойств
нервных и мышечных, свидетельствует о их крайне низкой электропроводности. Так
называемый аксон, диаметром 1 микрон и длиной 1 м, имеет сопротивление 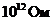 . Поэтому, в невозбудимой мембране
всякий сдвиг мембранного потенциала быстро затухает в окрестности того места,
где он возник, что полностью соответствует кабельным свойствам.
. Поэтому, в невозбудимой мембране
всякий сдвиг мембранного потенциала быстро затухает в окрестности того места,
где он возник, что полностью соответствует кабельным свойствам.
Возбудимым мембранам также присущ декремент потенциала, по мере удаления
от места возникновения возбуждения. Однако, если затухающий потенциал
достаточен для включения воротного процесса потенциал зависимых ионных каналов,
то на удалении от первичного очага возбуждения возникает новый ПД. Для этого
должно соблюдаться условие:
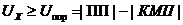
Регенерированный ПД также будет распределяться с декрементом, но, угасая
сам, он возбудит последующий участок волокна, и этот процесс повторяется
многократно:
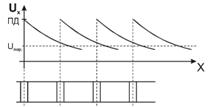
В силу огромной скорости декрементного распределения потенциала,
электроизмерительные приборы не способны зарегистрировать угасание каждого
предыдущего ПД на последующих участках БМ. Вдоль всей возбудимой мембраны, при
распределении по ней возбуждения, приборы регистрируют только одинаковые по
амплитуде ПД. Распределение возбуждения напоминает горение бикфордова шнура.
Создается впечатление, будто электрический потенциал распределяется по БМ без
декремента. На самом деле, бездекрементное движение ПД по возбудимой мембране,
является результатом взаимодействия двух процессов:
. Декрементного распределения потенциала от предыдущего ПД.
. Генерация нового ПД. Этот процесс называют регенерацией.
Первый из них протекает на несколько порядков быстрее, чем второй,
поэтому, скорость проведения возбуждения по волокну тем выше, чем реже
приходится ретранслировать (регенерировать) ПД, что, в свою очередь, зависит от
декремента потенциала вдоль БМ (). Волокно, обладающее большей , быстрее проводит нервные импульсы
(импульсы возбуждения).
В физиологии принят и иной подход для описания распределения возбуждения
по нервным и мышечным волокнам, которое не противоречит выше рассмотренному.
Этот подход был разработан Германном, и его называют методом локальных токов
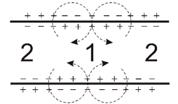
- возбудимый участок;
- невозбудимый участок.
Согласно этой теории, между возбудимым и невозбудимым участками волокна,
течет электрический ток, так как внутренняя поверхность первого из них обладает
положительным потенциалом относительно второго, и между ними существует
разность потенциалов. Токи, возникающие в живых тканях вследствие возбуждения,
называются локальными, так как распределяются на незначительное расстояние от
возбужденного участка. Их ослабление обусловлено затратами энергии на заряд
мембраны и на преодоление сопротивления цитоплазмы волокна. Локальный ток
служит раздражителем для покоящихся участков, которые непосредственно прилегают
к месту деполяризации (возбуждения). В них развивается возбуждение, а значит и
новая деполяризация. Она приводит к установлению разности потенциалов между
вновь деполяризированными и покоящимися (последующими) участками волокна,
вследствие чего, возникает локальный ток в следующем микроконтуре,
следовательно, распределение возбуждения представляет собой многократно
повторяющийся процесс.
. Факторы, влияющие на скорость распределения возбуждения
Скорость распределения возбуждения возрастает по мере понижения
сопротивления цитоплазмы и емкости клеточной мембраны, так как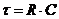 , сопротивление определяется по
формуле:
, сопротивление определяется по
формуле:
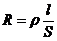
- длина нервного волокна;
- сечение нервного волокна;
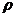 - удельное сопротивление цитоплазмы.
- удельное сопротивление цитоплазмы.
Толстые волокна обладают низким сопротивлением, и, вследствие этого,
быстрее проводят возбуждение. Так, в ходе эволюции, некоторые животные
приобрели способность к быстрой передаче нервных импульсов, за счет образование
в них толстых аксонов, путем слияния многих мелких в одно крупное. Примером
служит гигантское нервное волокно кальмара. Его диаметр достигает 1-2 мм, тогда,
как обычное нервное волокно имеет диаметр от 1-10 микрон.
Эволюция животного мира привела и к использованию другого пути повышения
скорости передачи нервной импульсации, то есть, уменьшению емкости
плазматической мембраны аксона (аксолемма). В результате появились нервные
волокна, покрытые миелиновой оболочкой. Они называются мякотными или
миелиновыми. Миелиновая оболочка образуется в процессе "наматывания"
на аксон клеток. Оболочка представляет собой много мембранную систему,
включающую от нескольких десятков, до 200 элементов клеточных мембран, которые
прилегают друг к другу и, при этом, внутренний их слой образует плотный
электрический контакт с аксолеммой. Толщина всей миелиновой оболочки
сравнительно невелика (1 микрон), но это достаточно для значительного понижения
емкости мембраны. Так как миелин является хорошим диэлектриком (удельное
сопротивление миелиновой оболочки составляет примерно 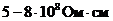 ), емкость мембраны миелинового
аксона примерно в 200 раз меньше емкости аксона без мякотного волокна, то есть,
примерно 0,005 и
), емкость мембраны миелинового
аксона примерно в 200 раз меньше емкости аксона без мякотного волокна, то есть,
примерно 0,005 и 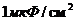 соответственно.
соответственно.
Диффузия ионов через миелиновую оболочку практически невозможна, кроме
того, в участках аксона, покрытых ею, отсутствует потенциал зависимые ионные
каналы. В этой связи, в мякотном нервном волокне, места генерации ПД
сосредоточены только там, где миелиновая оболочка отсутствует. Эти места в
мембране миелинового аксона называются перехватами Ранвье или активными узлами.
От перехвата к перехвату нервные импульсы проводятся за счет
декрементного распределения электромагнитного поля (движение локальных токов).
Расстояние между соседними перехватами составляет в среднем 1 мм, но оно
сильно зависит от диаметра аксона. Так, например, у животных эта зависимость
выражается следующим образом:
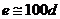
Перехваты Ранвье занимают примерно 0,02% общей длины нервного волокна.
Площадь каждого из них около 20 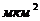 .
.
Время проведения возбуждения между соседними активными узлами составляет
примерно 5-10% длительности ПД. В этой связи, сравнительно большой путь (около
1 мм) между следующими друг за другом участками ретрансляции ПД обеспечивает
высокую скорость проведения нервного импульса. Необходимо отметить, что локальные
токи, достаточные для регенерации ПД, могут даже протекать через 2-3
последовательно расположенные перехвата Ранвье. Более частое, чем необходимо
для обеспечения нормального распределения возбуждения, расположение активных
узлов в мякотных аксонах, служит повышением надежности нервных коммуникаций в
организме. У гомойотерных животных надежность выше, чем у пойкилотерных
(животных с переменной температурой). В без-мякотных аксонах ретрансляция ПД
происходит значительно чаще. Там генераторы ПД расположены вдоль всей длины
волокна, в непосредственной близости друг от друга (около 1 микрона). Это
обусловлено сравнительно низкой скоростью проведения возбуждения по мембранам
мышечных и нервных волокон, которые не покрыты миелиновой оболочкой. В отличие
от них, миелиновые аксоны за счет малой емкости между участками перехватов
Ранвье, приобрели высокую скорость передачи нервных импульсов (до 140 м/с).
Вследствие относительно большой протяженности участков аксона между
соседними активными узлами, проведение нервного импульса в мякотном нервном
волокне, происходит как бы скачками, и поэтому его называют сальтоторным.
Сальтоторные проведения обеспечивают существенную экономию энергии. Так,
например, потребление при нем, в 200 раз меньше, чем при непрерывном распределении
нервных импульсов по без мякотным аксонам. Наибольшая скорость распределения
возбуждения наблюдается в мякотных аксонах, диаметр которых составляет примерно
10-15 микрон, а толщина миелиновой оболочки достигает 30-50 % общего диаметра
волокна. Скорость проведения нервных импульсов в миелиновых аксонах
пропорциональна их диаметру. Тогда, как в без мякотных аксонах, скорость
проведения возбуждения пропорциональна корню квадратному из диаметра.
. Биофизические основы электрокардиографии. Элементы структры миокарда и
их мембранные потенциалы
Механизмы биоэлектрогенеза в миокарде те же самые, что и в других
возбудимых тканях. Источником электромагнитной энергии служит концентрационный
элемент. ПП определяется более высокой проницаемостью плазмолеммы
миокардиального волокна (сарколеммы) для ионов по сравнению с высокомолекулярными
анионами, которые образуют соли в саркоплазме. ПД обусловлены открытием
натриевых потенциалзависимых каналов в сарколемме. Некоторый вклад в ПД
миокардиальных волокон, вносят ионы . Возбуждение распространяется по
сердцу без декремента, благодаря существованию в миокардиальных волокнах
положительной обратной связи между локальными токами и генерацией ПД. Вместе с
тем, биоэлектрогенез (БЭГ) в миокарде имеет ряд своеобразий.
Сердечная мышца неоднородна в своем клеточном составе. Различают типичные
(сократительные) миокардиокальные волокна (ТМВ) и атипичные миокардиокальные
волокна (АТМВ).
Между ними выявлены существенные различия в структуре, функции и
электрогенезе.
Мембранные потенциалы для АТМВ и ТМВ имеют следующий вид:
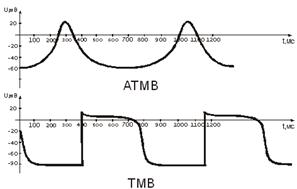
Мембранные потенциалы миокарда
Для типичных миокардиальных волокон (ТМВ), образующих основную массу
сердечной мышцы и осуществляющей ее сократительную деятельность, характерны
мембранные потенциалы (рис. Б). В них отчетливо просматривается стабильный
уровень ПП (~-90мВ). Возбуждение ТМВ проявляется в потенциале действия, восходящая
ветвь которого отображает изменение потенциала волокна (относительно
межклеточной среды) от - 90 до +10 (максимум до + 20 мВ), происходящее примерно
за 20 мс. В отличие от мышечных волокон и аксонов, миокардиальное волокно
обладает большим потенциалом инверсии (не более +20 мВ). Однако наиболее
характерные особенности присущи реполяризации сердечной мышцы. Если в волокнах
скелетной мышцы реполяризация происходит в течение короткого времени (около 10
мс), то репорялизия ТМВ продолжается 250-350 мс. На реполяризационной ветви ПД
ТМВ выражены три части:
- начальная быстрая реполяризация
- медленная реполяризация (плато)
конечная быстрая реполяризация.
Реполяризация завершается возвращением мембранного потенциала на
устойчивый уровень ПП.
Иначе выглядят мембранные потенциалы, регистрируемые в атипичных волокнах
миокарда (АТМВ). В них нет устойчивого уровня поляризации, то есть нет ПП.
Электрическая активность АТМВ представляет собой непрерывные колебания
мембранного потенциала. По достижении им определенной величины (примерно -60мВ)
спонтанно (произвольно) начинает развиваться медленная поляризация. Затем (на
уровне примерно -40 мВ) процесс ускоряется, что соответствует возникновению в
АТМВ ПД с присущей ему инверсией. Деполяризация сменяется реполяризацией, в
конце которой снова начинается медленная реполяризация. Следовательно для АТМВ
характерна безостановочная динамика мембранных потенциалов, лежащая в основе
самопроизвольной ритмической деятельности сердца.
Свойство миокарда возбуждаться под влиянием ПД, который спонтанно
возникает в нем самом (без внешних стимулов), называется автоматизмом
(автоматией). Биофизическая природа автоматизма пока не выявлена. Электрические
процессы, связанные с ним происходят в АТМВ. Установлено, что именно АТМВ служит
источником автоматизма сердечной мышцы.
. Механизм распространения возбуждения по миокарду
Ткань АТМВ сосредоточена в виде островков в различных отделах сердца. При
таком распространении, АТМВ объединяются в единую систему, когда по сердцу
происходит распространение возбуждения. Особенности проведения возбуждения по
миокарду связаны со способом соединения между собой его отдельных волокон. Под
электронным микроскопом между волокнами обнаружены так называемые вставочные
диски. Они представляют собой дубликаторы мембран, в которых имеются щелевые
контакты между волокнами, обеспечивающие электрическую синаптическую передачу.
Ранее считали, что будто "протоплазматические мостики" между
миокардиальными волокнами объединяют весь миокард в синцитий, под которым понимали
цитоплазмазматическую (проводящую) среду, не разделенную мембранами. Открытие
вставочных дисков внесло коррективу в морфологическое определение
миокардиального синцития, но не опровергло представления о миокарде, как
функциональном синцитии, поскольку электрический импульс (ПД), возникший в
одном из волокон, распространяется на соседние посредством электрической, а не
химической передачи. Благодаря этому сердцу присущи свойства отдельной клетки.
Необходимо подчеркнуть, что электрическая связь существует не только
между ТМВ сердца. Аналогично связаны АТМВ как между собой так и с ТМВ и
распространяется по всему сердцу.
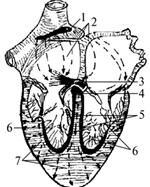
Рис. Схема "проводящей системы" сердца.
Обозначения: 1 - синсо-аурикулярный узел (узел Кейта-Флака); 2 - миокард
предсердий; 3 - атрио-вентрикулярный узел (узел Ашоффа-Тавары); 4 - пучок Гиса;
5 - ножки пучка Гиса; 6 - волокна Пуркинье; 7 - миокард желудочков. Стрелками
показан путь распространения возбуждения.
Между островками тканей АТМВ установлена четкая субординация: ведущим
является тот островок, который расположен в правом предсердии - между венозным
синусом и ушком (auricolus). От расположения произошло его название - сино -
аурикулярный узел. Когда он автоматически возбуждается (обычное состояние у
здорового человека), другие островки АТМВ выполняют только функцию проведения
возбуждения. При этом их собственный автоматизм подавляется. Поэтому сино -
аурикулярный узел называют пейсмекером (отводитель ритма). Ритм его
автоматических возбуждений определяет частоту сердечных сокращений.
Возбуждение спонтанно, в определенном ритме задается в сино -
аурикулярном узле. Каждое его АТМВ связано со своими соседями примерно сотней
щелевых контактов. Они обеспечивают взаимодействие между собой отдельных волокон
узла при становлении единичного сердечного ритма, а также обеспечивают
проведение возбуждения из узла в предсердие.
Благодаря тому, что водителем ритма является не одиночное АТМВ, а группа
из 5000 волокон, сердце имеет высокую надежность в поддержании стойкого периода
спонтанных колебаний. Экспериментально установлено, что в системе из-за АТМВ
период колебаний крайне не устойчив, изменяясь в пределах 200% (от 340 до 620
мс), тогда как в системе из 100 АТМВ его вариации не выходят за 20%, то есть
период изменяется от 500 до 580 мс.
С АТМВ сино-аурикулярного узла возбуждение переходит на ТМВ предсердий и
переводится по ним со скоростью примерно 1 м/с. Уже через 40 мс после
возникновения ПД в водителе ритма, все участки предсердий находятся в
возбужденном состоянии. Возбуждение распределяется по предсердиям с широким
фронтом (радиально), что обеспечивается наличием щелевых контактов не только
между торцами отдельных волокон, но и торцевыми контактами по их бокам. Однако,
плотность щелевых контактов в торцах вставочных дисков значительно выше, чем в
боковых. Вследствие этого постоянная длины (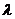 ) для продольного направления
составляет примерно 1 мм, а для поперечного в 3-10 раз меньше. В этой связи, по
направлению к желудочкам возбуждение движется быстрее, чем поперек предсердий.
Вместе с тем, существование торцевых вставочных дисков приводит к тому, что
возбуждение распределяется по предсердиям единым фронтом (радиально). Боковые
электрические связи выравнивают фронт, благодаря чему возбуждение, идущее в
предсердиях по разным путям, практически одновременно достигает второго
островка АТМВ - атрио-вентикулярного (предсердо-желудочкового) узла.
) для продольного направления
составляет примерно 1 мм, а для поперечного в 3-10 раз меньше. В этой связи, по
направлению к желудочкам возбуждение движется быстрее, чем поперек предсердий.
Вместе с тем, существование торцевых вставочных дисков приводит к тому, что
возбуждение распределяется по предсердиям единым фронтом (радиально). Боковые
электрические связи выравнивают фронт, благодаря чему возбуждение, идущее в
предсердиях по разным путям, практически одновременно достигает второго
островка АТМВ - атрио-вентикулярного (предсердо-желудочкового) узла.
Предсердия отделены от желудочков фиброзной тканью, которая не способна
проводить возбуждение, однако, в этой преграде есть узкая щель, шириной чуть
больше 1 мм и длиной примерно 1,5-2 мм. В этой щели и расположен
атрио-вентикулярный узел, который проводит возбуждения из предсердий в
желудочки. Другого пути для проведения возбуждения нет. В местах контакта с ТМВ
предсердий, АТМВ атрио-вентикулярного узла очень тонки, вследствие чего им
присуще значительное электрическое сопротивление саркоплазмы. В этом одна из
причин резкого (в 20-50 раз) замедления распределения возбуждения в
атрио-вентикулярном узле по сравнению с предсердиями. Другая причина замедления
заключается в том, что АТМВ в верхней части узла имеет не продольное, а
поперечное расположение. Следовательно, по направлению к желудочкам возбуждение
передается через боковые, а не торцевые щелевые контакты. Постоянная длины () для такого проведения составляет
примерно 95 мкм, что обусловлено снижением скорости распределения возбуждения
на порядок. Она составляет в верхней части атрио-вентикулярного узла примерно
0,02-0,05 м/с.
Замедление проведения возбуждения из предсердий в желудочки обеспечивает
чрезвычайно важную для нормальной работы функцию, то есть паузу между
сокращениями. Желудочки начинают сокращаться примерно через 0,1 с от начала
сокращений предсердий. Задержка нужна для того, чтобы вся кровь, накопленная в
предсердиях в диастолу, полностью перешла в желудочки до того момента, как они
начнут сокращаться.
Торцевые контакты, расположенные в атрио-вентикулярном узле
перпендикулярно основному направлению возбуждения, обеспечивают постоянную
длины () примерно 270 мкм, и они
обеспечивают сравнительно быстрое распространение потенциалов между отдельными
волокнами АТМВ в поперечном направлении. Это делает весь узел единой системой,
в которой возбуждение движется единым фронтом. Не обладай узел такой
структурой, то из разных его точек с различной временной задержкой выходило бы
несколько импульсов в нижестоящие отделы сердца.
Из атрио-вентикулярного узла возбуждение единым фронтом поступает в
следующее звено проводящей системы - атрио-вентикулярный пучок (пучок Гиса).
Там скорость проведения быстро возрастает, достигая 2-3 м/с. Увеличение
скорости обусловлено в основном утолщением АТМВ и повышением плотности щелевых
контактов во вставочных дисках. Плотность увеличивается примерно на 20
процентов их площади. Пучок Гиса лежит в толще миокардов желудочков, подобно
изолированному кабелю, при этом он не образует синопсов (электрических контактов)
с ТМВ желудочков. Ближе к верхушке сердца, от пучка Гиса отходят волокна
Пуркинье. Эти атипичные миокардные волокна вступают в контакт с ТМВ желудочков.
Волокна Пуркинье обладают наибольшим диаметром, по сравнению с другими
волокнами миокарда, и значительной постоянной длины (), которая в десятки раз больше длины
отдельных волокон АТМВ. В этой связи скорость проведения возбуждения ими
достигает 4-5 м/с.
Связь между волокнами Пуркинье и ТМВ желудочков осуществляется
многочисленными тонкими разветвлениями АТМВ. Отдельные такие волокна действуют
на ТМВ, как точечный источник тока, поэтому от точки контакта между ними
деполяризация (ПД) распределяется в виде сферической волны. Здесь нет условий
для распределения фронта возбуждения, в силу чего, невелика. В этой связи, в контакте
разветвленной сети волокон Пуркинье с желудочковыми ТМВ, осуществляется
передача возбуждения с замедлением. Задержка импульса достигает 30 мс, что
обеспечивает синхронизацию возбуждения всего сократительного миокарда
желудочков. Возбуждение с отдельных волокон Пуркинье переходит на огромное
число ТМВ одновременно. Благодаря чему, различные участки желудочков
сокращаются симфазно. Задержка в проведении возбуждения с АТМВ на ТМВ
проявляется в том, что скорость его распределения по миокарду желудочков в этот
момент составляет всего 0,4 м/с. По миокарду желудочков возбуждение движется с
верхушки к основанию, то есть в противоположном направлении, относительно движения
по пучку Гиса. Последними возбуждаются те участки желудочков, которые
расположены около предсердий.
. Интегральный электрический вектор сердца (ИЭВС)
Сложный путь распространения возбуждения по сердцу отображается в
электрокардиограмме (ЭКГ), которой свойственна весьма характерная и причудливая
форма.
Моделью генератора ЭКГ является электрический токовый диполь. Согласно
этой модели миокард, при распределении возбуждения по нему представляется
совокупностью множества токовых диполей, которые постоянно возникают и
исчезают, то есть ежемоментно возникают новые диполи и исчезают старые. В
результате, на поверхности сердечной мышцы создается сложная динамическая
мозаика электрических потенциалов. В этой связи бесполезно контролировать
поведение отдельного электрического диполя для характеристики сердечной
активности, а целесообразнее перейти от множества диполей к одному
результирующему.
Результирующий дипольный момент равен векторной сумме всех отдельных
токовых диполей. Результирующий дипольный момент миокарда получил название
интегрального электрического вектора сердца (ИЭВС). Ритмический характер
автоматизма водителя ритма, а также передача возбуждения посредством
электрических синопсов обусловливают синфазность возбудительного процесса в
миокардных волокнах. Поэтому ИЭВС имеют сравнительно большую амплитуду
(особенно, при деполяризации желудочков), чем создается высокий уровень
биопотенциалов, которые отображают сердечную деятельность даже на поверхности
тела.
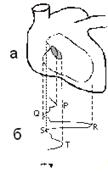
Типичные вектор электрокардиограммы (а) и электрокардиограммы (б)
В ходе сердечной деятельности ИЭВС претерпевают непрерывные изменения,
которые вполне закономерны, так как отображают закономерности распределения
возбуждения по миокарду. Ежемоментно амплитуда и направление ИЭВС - различны.
Измеряя их, врач получает информацию о движении волны возбуждения по сердцу.
Это позволяет ему оценить свойство миокарда и, в случае нарушений сердечной
деятельности, понять их причину.
При подаче электрических потенциалов, отображающих распределение
возбуждения по миокарду на пластины X и Y, на его экране возникают сложные
фигуры Лиссажу, которые образуются в результате сложения взаимно
перпендикулярных колебаний. В данном случае фигура Лиссажу представляет собой
траектории движения ИЭВС, которые описывает вектор на плоскости перпендикулярно
направлению распределения возбуждения (рис.1). Такое исследование электрической
активности сердца называется векторэлектрокардиоскопией (ВЭКС). На
векторэлектрокардиограмме выделяют обычно 3 эллипсоподобные фигуры. Самая малая
из них отображает деполяризацию предсердий, самая большая - деполяризацию
желудочков, и средняя - из реполяризацию.
Амплитуду вектора оценивают посредством измерения его проекций на
координатные оси. Любое из отведений ЭКГ есть ничто иное, как проекция ИЭВС. В
разнообразных способах отведения ЭКГ воплощаются различные системы координат.
Наиболее широкое распространение получила гексагональная система координат,
элементом которой является равносторонний треугольник. Эта система получила
название стандартных отведений. При ее практической реализации электроды
накладываются на левую ногу и запястья рук. Первым отведением считают
регистрацию разности потенциалов между двумя руками, вторым отведением - между
правой рукой и ногой, и третьим - между левой рукой и ногой.
. ЭКГ отведения как проекции ИЭВС на координатные оси
При любых отведениях биопотенциалов сердца от поверхности тела человека,
амплитуды зубцов ЭКГ представляют собой проекции ИЭВС на ту или иную ось
координатной системы в соответствующий момент сердечной деятельности. Зубец Р
отображает распределение возбуждения по предсердиям; комплекс QRS - при
возбуждении желудочков; зубец Т - при их реполяризации. Отклонение от нормы,
которое врач обнаруживает в том или ином элементе ЭКГ, дают ему информацию о
соответствующих процессах в той или иной части сердца.
Важнейшим параметром ЭКГ служат временные интервалы, по ним оценивают
скорость распределения возбуждения в каждом из отделов проводящей системы
сердца. Изменения скорости проведения связывают с повреждениями миокардных
волокон. Так, даже малый очаг поражения ТМВ диаметром 5-10 мкм, вызывает
задержку в распределении возбуждения на 0,1 мс.
В стандартных отведениях зубец Р обычно имеет амплитуду не более 0,25 мВ,
а его длительность равна 0,07-0,10 с. Интервал PQ отображает
атрио-вентикулярную задержку, и он составляет примерно 0,12-0,21 с при частоте
сердечных сокращений от 130 до 70 в минуту. Комплекс QRS наблюдается в течение
всего времени, пока возбуждение распределяется по желудочкам. Его длительность
изменяется в пределах от 0,06 до 0,09 с. Зубец Q в трети наблюдений отсутствует
в нормальной ЭКГ, а когда он обнаруживается, то его амплитуда не превышает 0,25
мВ. Зубец R обладает максимальной амплитудой среди всех других элементов ЭКГ, и
его амплитуда меняется в пределах 0,6-1,6 мВ. Зубец S также зачастую
отсутствует, но когда его обнаруживают, может иметь амплитуду до 0,6 мВ. Его
появление на ЭКГ характеризует тот процесс, когда возбуждение по миокарду
желудочков завершается вблизи основания (у предсердий). Интервал TS при пульсе
65-70 сокращений в минуту, составляет примерно 0,12 с. Длительность зубца Т
обычно меняется в пределах от 0,12 до 0,16 с, а его амплитуда изменяется в
пределах 0,25-0,6 мВ.
Необходимо отметить, что зубец Р возникает на ЭКГ примерно за 0,02 с до
начала сокращения предсердий, а комплекс QRS- за 0,04 с до начала сокращения
желудочков. Следовательно, электрические проявления возбуждения предшествуют
механическим (сократительные деятельности миокарда). В этой связи нельзя
говорить, будто ЭКГ является результатом сердечной деятельности (сердечные
сокращения). Имея ряд отведений ЭКГ (не менее двух), снятых в разных
отведениях, можно синтезировать ИЭВС. В медицинской литературе его называют
электрической осью сердца. По определению, электрическая ось сердца - это
отрезок прямой (вектор), соединяющий два сечения миокарда, обладающих в данный
момент наибольшей разностью потенциалов. Этот вектор направлен от
отрицательного полюса (возбужденного участка) к положительному (покоящемуся
участку). Направление электрической оси сердца в ходе распределения возбуждения
по миокарду, постоянно меняется, в этой связи принято определять среднюю ось
сердца. Так называют вектор, который можно построить в промежутках между
началом и окончанием деполяризации миокарда желудочков. По расположению средней
оси оценивают геометрическую ось сердца, которые, как правило, параллельны друг
другу. Таким образом, построенная средняя электрическая ось сердца дает
представление о положении сердца в грудной полости, и ее изменение служит
признаком в изменениях соответствующего желудочка.
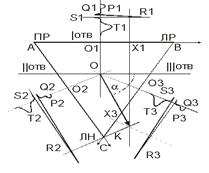
. Биомеханика кровообращения. Элементы биомеханики сердца
Кровообращение происходит в сердечно-сосудистой системе, которую
схематично можно представить в следующем виде:
Схема сердечно-сосудистой системы человека
С позиции механики, сердце является комбинированным насосом, который
нагнетает кровь в большой (1) и малый (2) круги кровообращения. Сосудистая
система образована сосудами разного типа: аорты, артерии, артериоллы,
капилляры, вены. Механическая работа сердца совершается за счет сократительной
деятельности миокарда. Вслед за распределением возбуждения происходит
сокращение миокардных волокон.
Вследствие ритмичного характера автоматизма, сердцу свойственна
цикличность сократительной деятельности. Каждый цикл сердечных сокращений
состоит из ряда механических процессов, идущих в строгой последовательности.
Необходимо отметить, что ЛЖ нагнетает кровь в большой круг кровообращения, а ПЖ
- в малый круг. При этом, ЛП совершает работу по наполнению кровью ЛЖ и тем
самым способствует эффективности его нагнетательной функции. То же самое делает
ПП по отношению к ПЖ. При этом, принято условно говорить о левом и правом
сердце, как о двух насосах, соединенных друг с другом последовательно. Выход
одного их них является входом для другого. Оба насоса перекачивают одинаковый
объем крови за один цикл.
Проанализируем цикл сердечной деятельности при частоте пульса 75
сокращений в минуту. В этом случае полное время сердечного цикла составляет 0,8
с. Механический цикл деятельности сердца можно представить в виде:
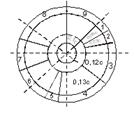
Цикл сердечной деятельности человека при пульсе 75 сокращений в минуту
9
-систола
предсердий;
1-4-систола желудочков;
-9-диастола желудочков
- фаза асинхронного сокращения;
- фаза изометрического сокращения;
- фаза быстрого изгнания крови;
- фаза медленного изгнания крови;
- протодиастолический интервал;
- фаза изометрического расслабления;
- фаза быстрого наполнения кровью;
- фаза медленного заполнения кровью;
- пресистола желудочков, систола предсердий.
Сначала сокращаются предсердия в течение 0,1 с. Благодаря
атрио-ветрикулярной задержке, их сокращение происходит при расслабленных
желудочках. В результате, вся кровь из предсердий перетекает в желудочки. Затем
начинают сокращаться желудочки. В желудочковой систоле выделяют ряд стадий
(фаз). Первая из них - фаза асинхронного сокращения, которая длится 0,05 с. В
течение этой фазы нет полной симфазности в возбуждении миокардных волокон.
Из-за неодинаковости (неодновременности), сокращения отдельных волокон в
желудочке развивают слабое усилие, но к концу этой фазы симфазность нарастает,
и уже многие волокна вовлекаются в сокращение, вследствие чего, развивается
достаточная сила, чтобы создать внутри желудочков большее кровяное давление КД,
чем в предсердиях (5 мм рт.ст. или 0,66 кПа). В этот момент КД в магистральных
артериальных стволах, куда кровь затем должна быть выброшена в желудочки,
значительно выше. Так, в аорте большой круг кровообращения примерно 60 мм
рт.ст. или 8 кПа, а в легочных артериях - примерно 15 мм рт.ст. или 2 кПа.
Каждый из желудочков отделен от своего предсердия створчатым клапаном, а
от соответствующего магистрального ствола - полулунным клапаном. Клапаны
открываются и закрываются в зависимости от соотношения КД в полостях, которые
ими разделяются. Створчатые клапаны закрыты тогда, когда КД внутри желудочков
выше, чем в предсердиях. Полулунные клапаны напротив, открываются при высоком
давлении в желудочках (при КД в ПЖ выше, чем в легочном стволе, и при КД в ЛЖ
выше, чем в аорте). Когда к концу фазы асинхронного сокращения внутрижелудочковое
КД станет выше, чем внутрипредсердное, створчатые клапаны мгновенно закроются.
При этом миокард продолжает сокращаться в условиях герметичности своих
полостей, которые заполнены кровью. Следовательно, первая фаза систолы
желудочков сменяется второй фазой - изометрическими сокращениями.
Изометрический режим обусловлен тем, что миокард сокращается вокруг
несжимаемого тела - крови, которая содержится в желудочках, и она не может
выйти из них, так как обе пары отверстий перекрыты клапанами (створчатый и
полулунный клапаны закрыты). Существование фазы изометрического сокращения
является важным условием для нормальной сердечной деятельности. Все усилие,
развиваемое миокардом, идет на повышение его потенциальной энергии, и,
следовательно, на создание напряжения. В результате чего, КД внутри Ж
стремительно нарастает: в ЛЖ от 5 до 60 мм рт.ст.; в ПЖ от 5 до 15 мм рт.ст.
это происходит за 0,03 с - такова продолжительность фазы изометрического
сокращения.
Благодаря достижению внутри Ж КД больше, чем в магистральных артериальных
стволах, открываются полулунные клапаны, и кровь устремляется из сердца в
сосуды. Вначале КД в Ж продолжает еще повышаться, несмотря на то, что
полулунные клапаны открыты. Этот режим сокращения называют изотермическим, что
не совсем точно выражает его характер, так как при нем КД не удерживается на
постоянном уровне. Так, например, в ЛЖ КД повышается примерно еще на 60 мм
рт.ст. за 0,12 с. За это время кровь быстро вытекает из Ж в сосудистое русло. В
этой связи третья фаза сердечного цикла называется фазой быстрого изгнания
крови. Затем еще 0,13 с кровь вытекает из Ж в фазу медленного изгнания. При
этом, в течение четвертой фазы, скорость истечения крови понижается, поскольку
падает внутрижелудочковое давление. Так, КД в ЛЖ, достигшее к началу фазы
медленного изгнания 120-130 мм рт.ст., к концу четвертой фазы уменьшается, и
становится меньше диастолического давления в аорте (меньше 60 мм рт.ст.). после
чего, полулунные клапаны закрываются, и наступает диастола желудочков, которая
также имеет несколько фаз. Таким образом, сердечный цикл повторяется
многократно.
. Работа сердца
Сила, возникающая в миокарде при его сокращении, зависит от исходной
длины миокардных волокон, поскольку, по мере укорочения, волокно увеличивает
число миозиновых мостиков, которые участвуют в сокращении. Это является общим
свойством мышечной ткани. При этом, исходная длина миокарда зависит от степени
растяжения их кровью, поступающей из предсердий. Следовательно, сила сокращения
миокарда желудочков тем значительнее, чем больше крови поступает в них во время
диастолы. От диастолического наполнения желудочков зависит основной показатель
сократительной функции сердца - систалический (ударный) объем - Vс - это, так
называемый объем крови, который выбрасывается каждым желудочком при одном
сокращении. Систалические объемы (Vс) ЛЖ и ПЖ одинаковы. Для каждого из
желудочков Vс составляет примерно 70-100 мл в условиях покоя. При этом
изгоняется примерно 60-70% крови, содержащейся в желудочках в период диастолы.
При физической работе и эмоциях Vс увеличивается в 2-3 раза. От Vс зависит
работа сердца, но ее величина может быть разной при одном и том же Vс.
Работа, совершаемая сердцем, затрачивается, во-первых, на выталкивание
крови в магистральные артериальные сосуды против сил давления и, во-вторых, на
придание крови кинетической энергии. Первый компонент работы называется
статическим (потенциальным); второй компонент - кинетическим.
Статический компонент работы сердца вычисляется по формуле:
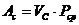
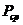 - среднее КД в соответствующем магистральном стволе (это
аорта для ЛЖ и легочная артерия для ПЖ).
- среднее КД в соответствующем магистральном стволе (это
аорта для ЛЖ и легочная артерия для ПЖ).
Поскольку изменение КД в артериях является сложной периодической функцией
от времени, среднее давление равно не полусумме максимального (систолического)
и минимального (диастолического), а среднему из множества малых изменений в
течение одного сердечного цикла. Среднее давление определяется по формуле:
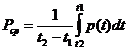
КД в артериальных стволах имеет вид:
Кривая динамики кровяного давления в артерии большого круга
кровообращения
Рс - систолическое давление; РД - диастолическое давление;
Рср - среднее давление; АД - время сердечного цикла
В отличии от систолического и диастолического давлений изменяющихся в
широких пределах, характеризуется постоянством. Величина в БКК составляет примерно 100 мм
рт.ст. (БКК - большой круг кровообращения) или 13,3 кПа, а в МКК Рср примерно
15 мм рт.ст. или 2 кПа, то есть в 6 раз меньше, чем в БКК. Однако, поскольку Vс
обоих желудочков одинаков, а давления, против которых они совершают работу,
имеют шестикратные различия, то и статический компонент ЛЖ в 6 раз больше статического
компонента ПЖ. Так, для ЛЖ статический компонент работы равен примерно 0,9 Дж,
а для ПЖ примерно 0,15 Дж, при Vс в 70 мл.
Кинетический компонент работы сердца определяется по формуле:
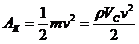
- плотность крови (близка к воде);
- скорость крови магистральной артерии (в состоянии покоя
равна 0,7 м/с). Следовательно, Ак = 0,02 Дж (при Vс = 70 мл).
В целом, работа ЛЖ за одно сокращение в условии покоя, составляет
примерно 1 Дж, а ПЖ - менее 0,2 Дж. При этом Ас доминирует над Ак и составляет
примерно 98% всей работы (а Ак примерно 2%). Средняя мощность миокарда в
условии покоя составляет примерно 1 Вт. При физической работе и эмоциях,
повышается КД в магистральных сосудах, и увеличивается Vс, но в еще большей
степени возрастает скорость кровотока. Следовательно, при физических и
психологических нагрузках вклад кинетического компонента в работу сердца может
достигать до 30% всей работы, а средняя мощность возрастает до 8 Вт.
. Основные показатели термодинамики
Движение крови по кровеносным сосудам изучает гемодинамика. Принято
выделять две группы показателей, которые интегрально характеризуют
кровообращение. Их называют гемодинамическими показателями. Первую группу
составляет скорость кровотока; вторую группу составляет КД. Различают объемную
и линейную скорость кровотока.
Объемной скоростью называют объем жидкости (крови), протекающей в единицу
времени (обозначают Q):
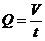 [мл/с] или [л/мин]
[мл/с] или [л/мин]
Линейная представляет собой путь проходимый частицами крови в единицу
времени (обозначают v):
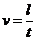 [м/с]
[м/с]
Связь между Q и v следующая:
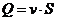 ,
,
где S - площадь поперечного сечения русла, по
которому течет кровь.
Для сплошного течение несжимаемой жидкости выполняется условие
неразрывности струи, то есть, через любое сечение струи в единицу времени,
протекают одинаковые объемы жидкости. Условие неразрывной струи выполняется и в
гемодинамике, где принята следующая формулировка: в любом сечении
сердечно-сосудистой системы объемная скорость кровотока одинакова, одной из
моделей круга кровообращения (как большого, так и малого) служит так называемая
разветвленная сосудистая трубка, которую схематично можно представить следующим
образом:
Схема разветвления сосудов в большом круге кровообращения
- аорта; 2 - магистральные артерии; 3 - крупные артерии
Каждое сечение разветвленной сосудистой трубки представляет собой
поперечный разрез всех кровеносных сосудов одного уровня ветвления. Так,
например, в БКК первое сечение проходит через аорту, второе - через все
магистральные артерии, на которые аорта непосредственно разветвляется, третье -
через все ветви артерий и т.д. В одном сечении находятся все капилляры БКК.
Площадь последнего сечения БКК (перед предсердиями) равна сумме поперечных
разрезов верхней и нижней полых вен.
Соотношение между суммарным поперечным сечением на разных уровнях
ветвления и линейной скоростью кровотока имеет следующий вид:
Соотношение между суммарным поперечным сечением сосудистой системы (S) на
разных уровнях ветвления и линейной скоростью (V) кровотока
- аорта; 2 - магистральные артерии; 3 - артериолы; 4 - капилляры; 5 -
вены
Из рисунка следует, что самым узким сечением в БКК является аорта.
Суммарная площадь обоих полых вен несколько больше. Самое обширное сечение
сосудистой трубки приходится на уровне капилляров. В БКК площадь их суммарного
просвета в 700-800 раз меньше, чем в аорте, и составляет примерно 1 мм/с. В
состоянии покоя средняя скорость в аорте лежит в пределах от 0,5 до 1 м/с, а
при больших физических нагрузках она может достигать 20 м/с. Кровь движется с
неодинаковой скоростью в разных участках сечения сосуда. Профиль скорости
кровотока имеет вид параболы, которую можно представить в виде:
Примерное распределение линейной скорости частиц крови в продольном
сечении кровеносного сосуда
При этом линейная скорость имеет максимальное значение на оси сосуда, и
постепенно уменьшается до нуля у его стенки. Пристеночный слой крови
неподвижен, следовательно, между слоями движущейся крови существует grad v ,
его называют скоростью сдвига и обозначают:
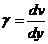 [1/с]
[1/с]
С grad v в поперечном профиле сосуда связана сила внутреннего трения
(вязкость) движущейся жидкости. Зависимость между ними выражается формулой
Ньютона:
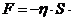
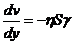
 - вязкость (коэффициент);
- вязкость (коэффициент);
- площадь поверхности, взаимодействующих слоев;
 - скорость сдвига.
- скорость сдвига.
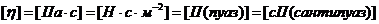
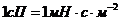
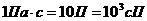
крови, измеряемая с помощью вискозиметра равна примерно 5 сП.
Это примерно в 5 раз больше, чем у воды и примерно такое же, как у
подсолнечного масла. В патологиях может изменяться от 1,7 до 23 сП.
Между вязкостью крови и воды обнаружена не только колличественная, но и
качественная разница. В воде, как и в других однородных жидкостях не зависит от . Такие жидкости называют
ньютоновскими. У крови принимает разное значение при разных . Так, 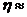 5 сП, что имеет место, при
сравнительно больших значениях, присущих движению крови в аорте. В других, более мелких
сосудах, оказывается значительно больше. Так,
в малых артериях 10 сП, а в капиллярах примерно 800 сП. Кровь вместе с другими
жидкостями, вязкость которых зависит от , относится к неньютоновским
жидкостям. Как правило, они представляют собой неоднородные жидкие среды (чаще
всего, суспензии). В крови зависимости () связана с наличием форменных
элементов (прежде всего, эритроцитов). При самых низких значениях (примерно 1
5 сП, что имеет место, при
сравнительно больших значениях, присущих движению крови в аорте. В других, более мелких
сосудах, оказывается значительно больше. Так,
в малых артериях 10 сП, а в капиллярах примерно 800 сП. Кровь вместе с другими
жидкостями, вязкость которых зависит от , относится к неньютоновским
жидкостям. Как правило, они представляют собой неоднородные жидкие среды (чаще
всего, суспензии). В крови зависимости () связана с наличием форменных
элементов (прежде всего, эритроцитов). При самых низких значениях (примерно 1 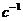 ), большая часть эритроцитов
объединяется в так называемые монетные столбики. Эти многоклеточные
конгломераты образуют жесткие структуры (твердое тело). Когда же превосходит предел текучести такого
твердого тела, монетные столбики вначале изгибаются, затем разрушаются.
Вследствие чего, кровь движется достаточно быстро, понижая свою вязкость. При
этом, тем в большей степени, чем выше скорость кровотока. При атеросклерозе крови возрастает, что приводит к
увеличению нагрузки на сердце.
), большая часть эритроцитов
объединяется в так называемые монетные столбики. Эти многоклеточные
конгломераты образуют жесткие структуры (твердое тело). Когда же превосходит предел текучести такого
твердого тела, монетные столбики вначале изгибаются, затем разрушаются.
Вследствие чего, кровь движется достаточно быстро, понижая свою вязкость. При
этом, тем в большей степени, чем выше скорость кровотока. При атеросклерозе крови возрастает, что приводит к
увеличению нагрузки на сердце.
От крови зависит характер ее течения по
сосудам, которое может быть:
·
ламинарным;
·
турбулентным.
Если слои крови движутся параллельно друг другу, не смешиваясь между
собой, то такое течение называют ламинарным. Когда поток жидкости
сопровождается перемешиванием слоев, то говорят о турбулентном (вихревом)
движении потока. Переход одного вида течения в другой определяется числом
Рейнольдса, которое равно:
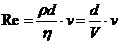
- плотность крови;
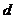 - диаметр сосуда;
- диаметр сосуда;
- вязкость крови;
- кинематическая вязкость;
- скорость.
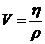
Если 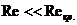 , то ламинарное, если
, то ламинарное, если  , то турбулентное.
, то турбулентное.
Для однородной жидкости Rе 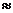 2300 для крови Rе = 970
2300 для крови Rе = 970 80.
80.
Ламинарное течение крови создает меньшую нагрузку на сердце, поскольку
при нем работа сердца пропорциональна объемной скорости в первой степени. При
турбулентном движении крови работа сердца пропорциональна Q в степени 1,7. В
нормальных условиях кровообращение преимущественно носит ламинарный характер.
При тяжелой физической нагрузке кровоток может перейти в турбулентный, в
основном, из-за повышения скорости кровотока (Q).
. КД. Гемодинамическое сопротивление
крови оказывает существенное влияние и на второй основной
гемодинамический показатель (КД). Для анализа факторов, от которых зависит КД,
рассмотрим уравнение Пуазеля относительно изменения давления, которое имеет
вид:

- длина кровеносного сосуда, соответствующего уровня ветвления;- радиус
сосуда соответствующего уровня ветвления;- объемная скорость кровотока.
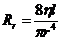 - гемодинамическое сопротивление.
- гемодинамическое сопротивление.
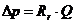
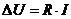
[ ]=[
]=[ ]
]
Величина Rr отображает сопротивление сосудистого русла кровотоку, включая
все факторы, от которых оно зависит. В этой связи Rr оказалось полезной
величиной для расчета и моделирования сердечно-сосудистой системы, при создании
искусственных аппаратов кровообращения, и при протезировании сердца и сосудов.
Характер уравнения Пуазелля относительно  свидетельствует, что КД зависит от
объемной скорости кровотока и следовательно, от массы циркулирующей крови и
сократительной деятельности миокарда, которые и определяют эту скорость. Еще в
большей степени оказывают влияние на динамику КД гемодинамическое сопротивление
(Rr) и, прежде всего, такой его компонент, как радиус сосуда, который входит в
формулу в 4-ой степени (
свидетельствует, что КД зависит от
объемной скорости кровотока и следовательно, от массы циркулирующей крови и
сократительной деятельности миокарда, которые и определяют эту скорость. Еще в
большей степени оказывают влияние на динамику КД гемодинамическое сопротивление
(Rr) и, прежде всего, такой его компонент, как радиус сосуда, который входит в
формулу в 4-ой степени ( ). В этой связи, изменение r на 20% вызывает изменение КД в
сосуде более чем в 2 раза. Таким образом, даже небольшие колебания просвета
кровеносных сосудов оказывают сильное влияние на кровообращение. Не случайно
регуляция уровня КД в организме связана с нервными или гуморальным
(химическими) влияниями, прежде всего, на гладкомышечную оболочку кровеносных
сосудов, в целях активного изменения их просветов. Как правило, на это же
направляются основные фармакологические средства, нормализующие КД.
). В этой связи, изменение r на 20% вызывает изменение КД в
сосуде более чем в 2 раза. Таким образом, даже небольшие колебания просвета
кровеносных сосудов оказывают сильное влияние на кровообращение. Не случайно
регуляция уровня КД в организме связана с нервными или гуморальным
(химическими) влияниями, прежде всего, на гладкомышечную оболочку кровеносных
сосудов, в целях активного изменения их просветов. Как правило, на это же
направляются основные фармакологические средства, нормализующие КД.
Проанализируем изменение Rr в системе кровообращения на модели
разветвленной сосудистой трубки. Rr в разных отделах сосудистой трубки имеет
следующее значение:
Гемодинамическое сопротивление в разных сечениях кровеносного русла
- аорта; 2 - артерии; 3 - артериоллы; 4 - капилляры; 5 - вены.
Все крупные артерии имеют большой радиус, который мало изменяется в
обычных условиях. Поэтому их вклад в Rr и в его изменение незначителен, хотя
длина артерии сравнительно велика. По мере удаление от желудочков, число
артерий, включаемых параллельно кровотоку возрастает, поскольку при
параллельном включении в кровообращение многих (n) сосудов Rr начинает падать в
n раз, и вклад этого звена артериального русла должен быть, казалось бы,
меньше, по сравнению с вкладом крупных артерий, тем более что по мере удаления
от сердца, каждое разветвление артерий становится короче. Однако, по мере
ветвления артерий, уменьшается их радиус, а, поскольку Rr зависит от r в 4
степени, то Rr становится тем больше, чем дальше от сердца расположено данное
артериальное русло. Особенно резкое увеличение Rr наблюдается на уровне
артериолл. Переход от артериолл к капиллярам характеризуется значительным
увеличением количества параллельно включаемых сосудов. Тогда, как радиус
прекапилляр (сосуда, принадлежащего последнему звену артериолл) и капилляра
примерно одинаковы. В этой связи общее Rr капиллярной сети примерно в 4 раза
меньше, чем артериолл. В венозном русле Rr еще меньше, чем в капиллярном.
Отмеченные особенности Rr в различных звеньях кровеносного русла
определяют КД в сердечно-сосудистой системе человека. Распределение КД в
различных сосудах БКК имеет следующий вид:
Распределение кровяного давления в различных сосудах большого круга
кровообращения человека
- аорта; 2 - крупные магистральные артерии; 3 - мелкие магистральные
артерии;
- артериоллы; 5 - капилляры; 6 - венулы; 7 - вены; 8 - полые вены.
В аорте и крупных артериях падение давления невелико. КД в начале и конце
таких сосудов почти одинаково. В артериоллах наблюдается максимальный перепад
давления, то есть, свыше половины всего общего падения КД в сосудистом русле. В
крупных и средних артериях КД неодинаково в систолу и в диастолу. В полых венах
КД имеет отрицательное значение. Смысл этого выражения состоит в том, что
давление крови в полых венах ниже атмосферного давления на несколько мм рт.ст.
принято различать следующие виды КД:
. Систолическое (max).
. Диастолическое (min).
. Пульсовое, равное разности максимального и минимального.
. Среднее.
Названные виды КД составляют вторую группу основных гемодинамических
показателей КД, то есть, той силы, с которой движущаяся кровь давит на единицу
поверхности соответствующих сосудов.
Функциональное предназначение кровеносных сосудов в системе
кровообращения неодинаково. В биофизике и физиологии принято различать 4 типа
кровеносных сосудов:
- Артерии эластичного типа. 2 - Артерии мышечного типа.
-Капилляры. 4- Вены.
1. Артерии эластичного типа поддерживают
кровоток во время диастолы желудочков сердца и, тем самым, обеспечивают
непрерывность движения крови в сосудистой системе.
2. Артерии мышечного типа создают
переменное сопротивление кровотоку и, следовательно, регулируют уровень КД в
системе кровообращения, а также объемную скорость кровотока в каждом из
органов.
3. Капилляры осуществляют обмен веществ
между кровью и тканями, ради чего и существует вся сердечно-сосудистая система.
4. Вены являются емкостными сосудами,
так как они обладают выраженной пластичностью и могут сильно деформироваться
(растягиваться) без существенного развития напряжения в них. В этой связи в
венах сосредоточено примерно 80% крови, находящейся в БКК.
. Элементы биофизики дыхания. Этапы механики дыхания
Газообмен между альвеолярной газовой смесью и атмосферным воздухом,
обеспечивающим диффузию 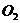 и
и 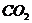 через альвеолярно-капиллярную мембрану (АКМ) осуществляется
благодаря работе аппарата вентиляции. Этот аппарат состоит из двух
анатомо-физиологических образований:
через альвеолярно-капиллярную мембрану (АКМ) осуществляется
благодаря работе аппарата вентиляции. Этот аппарат состоит из двух
анатомо-физиологических образований:
1. Грудной клетки с дыхательными
мышцами.
2. Легкими с дыхательными путями (при
сокращении).
Грудная клетка представляет собой жесткий подвижный футляр для легких
сердца и сосудов, который обладает упругостью. Грудная клетка активно изменяет
свой объем посредством сокращения диафрагмы и других дыхательных мышц. При
сокращении диафрагмы ее купол уплощается и смещается в сторону брюшной полости,
что приводит к увеличению объема грудной клетки, а, вслед за этим, в легких,
согласно закону Бойля-Мариотта, увеличение объема легких неизбежно
сопровождается понижением давления внутри них. В результате, давление
становится ниже атмосферного, и воздух засасывается внутрь легких (альвеол).
Таким образом совершается вдох. При спокойном дыхании 2/3 глубины вдоха
обеспечивается сокращением диафрагмы, а 1/3 - сокращением наружных межреберных
мышц. Усиление дыхания связано с включением в работу вспомогательных
дыхательных мышц (мышц шеи и плечевого пояса). Они могут способствовать
форсированию как вдоха, так и выдоха. В форсированный вдох наибольший вклад
вносят внутренние межреберные мышцы и мышцы живота (брюшной пресс).
При спокойном дыхании сокращаются дыхательные мышцы, обеспечивающие только
вдох, тогда, как выдох совершается пассивно, за счет возникновения при вдохе
силы упругости, как в легких, так и в тканях грудной клетки. Основные этапы
механики дыхания можно пояснить следующей схемой:
Этап вдоха.
1. Поступление первого импульса к дыхательным мышцам.
2. Синаптическая передача нервных импульсов.
. Увеличение объема грудной полости.
. Увеличение объема легких.
. Снижение давления воздуха в легких.
. Всасывание воздуха из атмосферы в легкие.
Этап выдоха.
1. Расслабление дыхательных мышц вслед за сокращением их при вдохе.
2. Уменьшение объема грудной полости.
. Уменьшение объема легких.
. Повышение давления в легких.
. Выдавливание воздуха из легких в атмосферу.
Энергия, затрачиваемая мускулатурой для осуществления свободного дыхания
равна примерно 2-2,5 Дж, при этом 70% расходуется на преодоление эластичного
сопротивления грудной клетки и легочной ткани, а остальные 30% идут на
совершение работы по перемещению воздуха против сил трения. Силы трения
пропорциональны скорости перемещения воздушной массы, следовательно второй
компонент энергозатрат возрастает при учащении дыхания. Энергозатраты на
преодоление эластичности органов дыхания практически не зависят от частоты
дыхания. В то время как энергозатраты на эластичное сопротивление в основном определяются
объемом воздуха, поступающего в легкие при вдохе, то есть глубиной дыхания.
Неодинаковая зависимость каждого из компонентов работы дыхательного аппарата от
частоты и глубины дыхания приводит к установлению оптимального соотношения
между ними. При этом соотношении работа должна поддерживаться на минимальном
уровне, при данном объеме вентиляции. Под объемом вентиляции понимают минутный
объем дыхания. В условиях физиологического покоя глубина дыхания обычно
составляет примерно 0,5 л, а частота 12-16 вдохов в минуту.
. Физические основы газообмена
В легких совершается обмен  между организмом и окружающей
средой. Благодаря работе дыхательных мышц легочные альвеолы обмениваются этими
газами с атмосферой. Это обеспечивает относительное постоянство и в альвеолярной газовой смеси.
Газообмен между альвеолярной газовой смесью и кровью легочных капилляров
происходит на альвеолярно-капиллярной мембране (АКМ). Схематично АКМ
представляется:
между организмом и окружающей
средой. Благодаря работе дыхательных мышц легочные альвеолы обмениваются этими
газами с атмосферой. Это обеспечивает относительное постоянство и в альвеолярной газовой смеси.
Газообмен между альвеолярной газовой смесью и кровью легочных капилляров
происходит на альвеолярно-капиллярной мембране (АКМ). Схематично АКМ
представляется:
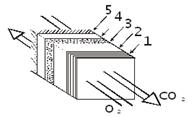
Схема альвеолярно-капиллярной мембраны и диффузии газов в легких
- альвеолярный эпителий, 2 - межклеточная среда, 3 - эндотелий капилляра,
4 - плазма крови, 5 - эритроцитарная мембрана
Газообмен в легких осуществляется под действием только одной силы -
концентрационного градиента каждого из газов на АКМ. Массоперенос газов
подчиняется уравнению Фика:
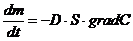
Используя уравнение Фика для количественной оценки газопереноса через АКМ
целесообразно заменить градиент концентраций на градиент парциальных давлений и
напряжений газов.
Парциальное давление газа - такое давление компонента газовой смеси,
которое он оказал бы на заключающую его оболочку, если бы один занимал весь
объем, предоставленный этой смеси.
Содержание газа в жидкости принято характеризовать напряжением. Под
напряжением газа в жидкости понимают такое парциальное давление данного газа,
которое нужно создать для поддержания равновесия в его содержании между газовой
смесью и жидкостью. То есть для прекращения всякого газообмена между ними.
Размерность парциального давления и напряжения - единица давления (Па -
мм.рт.ст.).
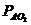 -парциальное давление со стороны альвеол.
-парциальное давление со стороны альвеол.
 -напряжение в капиллярах (в крови).
-напряжение в капиллярах (в крови).
Рассмотрим величины парциальных давлений и  в альвеолах газовой смеси. С одной
стороны и объем в крови легочных капилляров с другой. Это необходимо для оценки
сил под действием которых совершается обмен через АКМ. В газовой смеси легочных
альвеол
в альвеолах газовой смеси. С одной
стороны и объем в крови легочных капилляров с другой. Это необходимо для оценки
сил под действием которых совершается обмен через АКМ. В газовой смеси легочных
альвеол 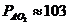 мм.рт.ст. или 13,7 кПа.
мм.рт.ст. или 13,7 кПа. 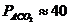 мм.рт.ст. или 5,3 кПа. Расчет
парциальных давлений в газовой смеси альвеол обычно производят по формуле:
мм.рт.ст. или 5,3 кПа. Расчет
парциальных давлений в газовой смеси альвеол обычно производят по формуле:
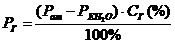
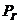 -парциальное давление газа.
-парциальное давление газа.
 -давление насыщенного газа паров воды в альвеолах при t=36,7
-давление насыщенного газа паров воды в альвеолах при t=36,7 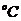 , =47 мм.рт.ст. или 6,3 кПа.
, =47 мм.рт.ст. или 6,3 кПа.
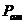 -атмосферное давление.
-атмосферное давление.
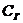 -концентрация данного газа в газовой смеси в %.
-концентрация данного газа в газовой смеси в %.
Парциальному давлению и в альвеолах газовой смеси противодействует на АКМ напряжение
этих газов в венозной крови легочных капилляров как 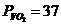 мм.рт.ст. или 4,9 кПа,
мм.рт.ст. или 4,9 кПа, 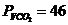 мм.рт.ст. или 6,1 кПа . Из
сопоставления и
мм.рт.ст. или 6,1 кПа . Из
сопоставления и 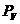 каждого из газов следует, что и преодолевают АКМ по концентрационным
градиентам, но в противоположных направлениях: из альвеолярной газовой смеси в
кровь, а наоборот.
каждого из газов следует, что и преодолевают АКМ по концентрационным
градиентам, но в противоположных направлениях: из альвеолярной газовой смеси в
кровь, а наоборот.
. Расчет газообмена
Производится расчет не массы газа, переносимого через АКМ, а его объем.
Тогда уравнение:
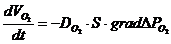
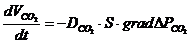
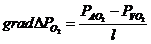

-длина, толщина АКМ.
Для полной оценки grad на АКМ необходимо знать толщину . АКМ состоит из двух слоев клеток:
·
эндотелия
легочных капилляров,
·
альвеолярного
эпителия
и базилярной мембраны, расположенной между ними. Общая толщина АКМ
примерно 0,2 мкм, но в области ядерных клеток примерно 10 мкм.
Кроме перечисленных компонентов эта многомембранная система, через
которую диффундирует газ. Легкие также включают пленку субфактата.
Основой субфактата является биомолекулярный липидный слой, подобный
фосфолипидному слою биологической мембраны.
Наслоение субфактата на эпителиальную поверхность альвеол создает
дополнительную к АКМ диффузионную среду, которую газы преодолевают при их
массопереносе. За счет субфактата удлиняется диффузионный путь, что в свою
очередь приводит к некоторому снижению концентрационных градиентов на АКМ.
Однако без субфактата дыхание вообще было бы невозможным, так как стенки
альвеол слиплись бы под действием значительного поверхностного натяжения,
которое присуще альвеолярному эпителию. Субфактат снижает поверхностное натяжение
альвеолярных стенок и предотвращает слипание альвеол.
Концентрационные градиенты и на АКМ резко падают при патологии.
Они могут уменьшатся как при изменении разности концентраций в альвеолах или
легочных капилляров, так и вследствие утончения АКМ. Первая причина характерна
для изменения газовой среды, в которой находится организм (при пребывании
человека в разряженной атмосфере), при нарушении легочного дыхания (при многих болезнях
легких, при недостаточной работе дыхательных мышц). Вторая патология - отек
легких. При этом жидкость, выходящая из капилляров скапливается в межклеточной
среде (базилярная мембрана). Это приводит к утоньшению АКМ и резко снижает
градиент концентрации газов на АКМ даже при нормальной разности их
концентрации.
В нормальных условиях диффузионный путь составляет примерно 5 мкм. На
этом расстоянии перепад концентраций и неодинаков. Он различается более чем
на порядок и выражается величинами парциального давления, составляющие
соответственно: для примерно 70 мм. рт. ст. или 9,3 кПа, для примерно 6 мм. рт. ст. или 0,8 кПа.
Но при столь значительных перепадах парциальных давлений и массоперенос этих газов одинаков 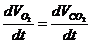 .
.
Причина того заключается в том, что и имеют разную проницаемость через
АКМ. Проникающая способность каждого из них оценивается коэффициентом диффузии.
Экспериментально установлено, что в воде 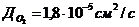 . В воздухе на 4-5 порядков выше (
. В воздухе на 4-5 порядков выше (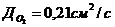 ). В липиде
). В липиде 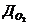 на несколько порядков ниже по
сравнению с водой. Проникающая способность через АКМ характеризуется
на несколько порядков ниже по
сравнению с водой. Проникающая способность через АКМ характеризуется 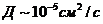 . Это позволило заключить, что проходит через АКМ сквозь поры,
заполненные водой. Также проходит через АКМ и , но он делает это в 20-25 раз
быстрее, чем , так как лучше растворяется в воде. Высокая степень растворимости в воде обуславливается его малым
(близким к нулю) дипольным моментом. Молекулы не поляризованы благодаря своей
симметричной структуре. Молекула не взаимодействует с заряженными
группами компонентов биологической мембраны и легко проходит сквозь нее. В этом
отношении с высочайшей проникающей способностью связано его участие во многих
процессах которые обеспечивают регуляцию дыхания, кровообращения, обмена
веществ и других функций организма.
. Это позволило заключить, что проходит через АКМ сквозь поры,
заполненные водой. Также проходит через АКМ и , но он делает это в 20-25 раз
быстрее, чем , так как лучше растворяется в воде. Высокая степень растворимости в воде обуславливается его малым
(близким к нулю) дипольным моментом. Молекулы не поляризованы благодаря своей
симметричной структуре. Молекула не взаимодействует с заряженными
группами компонентов биологической мембраны и легко проходит сквозь нее. В этом
отношении с высочайшей проникающей способностью связано его участие во многих
процессах которые обеспечивают регуляцию дыхания, кровообращения, обмена
веществ и других функций организма.
52. Диффузионная способность легких. Транспорт и
Для оценки газообмена в легких достаточно трудно измерить коэффициент
диффузии, так как для его расчета нужно знать величину площади диффузии,
которую невозможно измерить в каждом конкретном случае, чаще при ее оценке
ориентируются на площадь альвеолярной поверхности. В легких находиться
приблизительно … альвеол, каждая из которых имеет диаметр от 75 до 300 мкм. При
спокойном дыхании объем альвеол в среднем изменяется на 2-3%. При этих условиях
полная альвеолярная поверхность достигает 70-80 . Но площадь через которую газы
диффундируют определяется не всей площадью дыхательной поверхности альвеол, а
только той частью, где есть контакт с капиллярами, наполненными движущейся
кровью. Кровь текущая по легочным капиллярам создает тонкую пленку толщиной 10
мкм. Необходимо рассчитать поверхность так называемой кровяной плоскости, под
которой понимают суммарную поверхность эритроцитов, контактирующими со всеми
функционирующими альвеолами в течение одной минуты. Ежеминутно в состоянии
покоя через легкие протекает примерно пять литров крови. В них содержится
. Но площадь через которую газы
диффундируют определяется не всей площадью дыхательной поверхности альвеол, а
только той частью, где есть контакт с капиллярами, наполненными движущейся
кровью. Кровь текущая по легочным капиллярам создает тонкую пленку толщиной 10
мкм. Необходимо рассчитать поверхность так называемой кровяной плоскости, под
которой понимают суммарную поверхность эритроцитов, контактирующими со всеми
функционирующими альвеолами в течение одной минуты. Ежеминутно в состоянии
покоя через легкие протекает примерно пять литров крови. В них содержится 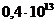 эритроцитов. Поверхность каждого из
них
эритроцитов. Поверхность каждого из
них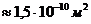 . Следовательно в состоянии покоя
площадь минутного газообмена в легких человека составляет примерно 3750. При физических нагрузках площадь
увеличивается до 12000-15000 .
. Следовательно в состоянии покоя
площадь минутного газообмена в легких человека составляет примерно 3750. При физических нагрузках площадь
увеличивается до 12000-15000 .
Благодаря значительной поверхности газообмена, полный массоперенос и в легких человека осуществляется
всего за 0,1 с. Вместе с тем, известно, что в условиях покоя каждый из
эритроцитов проходит легочный капилляр от начала до конца за примерно 2с. При физической
нагрузке скорость кровотока возрастает, что приводит, с одной стороны к
увеличению поверхности газообмена, приходящемуся на единицу времени контакта
крови и альвеолярной газовой смеси, а с другой стороны, к уменьшению
продолжительности этого контакта. Согласно закону Фика, сокращение времени
газообмена должно уменьшать объем газов диффундирующих через АКМ. Однако, это
утверждение справедливо только для очень кратковременного (короче 0,1 с)
газообмена. Когда же время кровотока по легочным капиллярам сокращается от 2 с.
(в состоянии покоя) до 0,1 с. (в состоянии нагрузки), массоперенос газов в
легких не уменьшается, а только увеличивается.
Приводимые выше аргументы позволяют понять насколько трудно оценить
влияние на газообмен человека таких параметров, как диффузионная площадь S и
время газообмена. В этой связи вводится своеобразная физическая величина,
которую называют диффузионной способностью легких -  Под легких понимают объем данного газа,
переносимого через АКМ всей дышащей поверхностью в течение 1 мин, при разности
парциальных давлений на мембране в 1 мм рт.ст. и нормальной толщине АКМ
(примерно 5 мкм). Другими словами, дыхательная способность ДЛ определяется
произведением коэффициента диффузии на площадь газообмена. ДЛ удается измерить
у человека и животных при помощи специальных приборов. В медицине принята
внесистемная единица измерения ДЛ =
Под легких понимают объем данного газа,
переносимого через АКМ всей дышащей поверхностью в течение 1 мин, при разности
парциальных давлений на мембране в 1 мм рт.ст. и нормальной толщине АКМ
(примерно 5 мкм). Другими словами, дыхательная способность ДЛ определяется
произведением коэффициента диффузии на площадь газообмена. ДЛ удается измерить
у человека и животных при помощи специальных приборов. В медицине принята
внесистемная единица измерения ДЛ =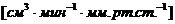
Зная величину ДЛ и grad парциальных давлений для и , можно вычислить газоперенос каждого
из газов через АКМ за 1 минуту:
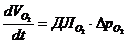
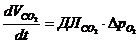
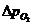 - разность между парциальным давлением и напряжением ;
- разность между парциальным давлением и напряжением ;
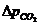 - разность между напряжением и парциальным давлением.
- разность между напряжением и парциальным давлением.
При изменении площади диффузии S, значение ДЛ также меняется. Так,
например, в покое 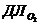 примерно 20-25,
примерно 20-25, 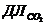 примерно 600 . При тяжелой физической
работе достигает примерно 70, а примерно 1700.
примерно 600 . При тяжелой физической
работе достигает примерно 70, а примерно 1700.
Проникнув в кровь, текущую по легочным капиллярам, переносится ею во все ткани
организма. Формы этого транспорта различны. Во-первых, растворяется в крови и, во-вторых, связывается с гемоглобином, который
содержится в эритроцитах. Наилучшей растворимостью в крови обладает , а растворяется меньше.
Коэффициент растворимости газов существенно зависит от свойств
растворителя. В крови газы растворяются хуже, чем в воде. Ухудшение
растворимости объясняется главным образом тем, что кровь содержит соли и белки.
Зная коэффициент растворимости в цельной крови при t примерно 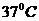 , а также напряжения в крови, оттекающей от легких
(примерно 100 мм рт.ст. или 13,3 кПа), легко можно рассчитать объем , растворяющегося в единице объема
артериальной крови. Подставив в уравнение Генри соответствующие значения, можно
убедиться, что в 100 мл крови растворяется 0,3
, а также напряжения в крови, оттекающей от легких
(примерно 100 мм рт.ст. или 13,3 кПа), легко можно рассчитать объем , растворяющегося в единице объема
артериальной крови. Подставив в уравнение Генри соответствующие значения, можно
убедиться, что в 100 мл крови растворяется 0,3 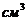 :
:

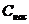 - концентрация газа;
- концентрация газа;
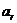 - коэффициент растворимости газа.
- коэффициент растворимости газа.
Однако, экспериментально установлено, что в 100 мл артериальной крови
примерно растворяется 20 . Различие обусловлено присутствием в эритроцитах
гемоглобина, каждый грамм которого связывает и переносит примерно 1,39 .
В каждых 100 мл крови обычно содержится 14-16 граммов гемоглобина, чем и
определяется кислородная емкость крови, то есть, объем транспортируется децилитром крови
(100 мл). При содержании гемоглобина в крови 14 г, кислородная емкость примерно
19 , а при 16 г гемоглобина - примерно
22 .
лучше растворяется в крови, в силу чего его объем
растворяемый в 100 мл крови составляет примерно 2,4 . В такой форме транспортируется
примерно 5% всего , переносимого кровью. Кроме этого, 15% кровь переносит в виде
карбамино-гемоглобина (соединения с гемоглобином), но наибольшая часть
(примерно 80%) транспортируется
кровью в виде бикарбонатов.
Азот переносится кровью только в растворенном состоянии, причем, в каждых
100 мл содержится всего 1 мг азота. Несмотря на столь ничтожное количество, все
ткани организма насыщены азотом, так как молекулы азота не участвуют в
метаболических процессах.
53. Элементы биофизики органов чувств
Организм представляет собой сложную самоорганизующуюся систему, которая
характеризуется обменом веществ, энергией и информацией с окружающей средой.
Целесообразное реагирование организма, при постоянно меняющихся условиях
внешней среды, возможно только при наличии непрерывного поступления в организм
информации от окружающей среды. Функцию получения и переработки информации об
условиях внешней среды в организме выполняют органы чувств. По существу, органы
чувств представляют собой измерительные устройства для анализа внешних
физических стимулов, а также для оценки эффективности действий, производимых
организмом. Таким образом, органы чувств выполняют роль обратной информационной
связи в системе "организм-среда". В 1968 г. Мэчи предложил следующую
схему, показывающую взаимодействие организма с внешней средой. Схема имеет вид:
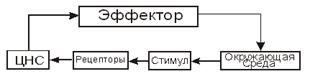
Схема, показывающая взаимодействие организма и внешней среды (по Мэчину)
При исследовании работы органов чувств представляют интерес два аспекта:
·
кибернетический;
·
биофизический.
Кибернетический аспект заключается в изучении принципов кодирования и
передачи информации в органах чувств.
Биофизический аспект заключается в исследовании конкретных
физико-химических процессов взаимодействия факторов внешней среды с органами
чувств, что приводит к трансформации энергии внешнего воздействия в
специфические сигналы, пригодные для анализа нервной системой.
При действии внешнего стимула на органы чувств у человека, возникает
специфическое ощущение. Зависимость между величиной ощущения и величиной
действующего стимула сформулирована в законе Вебера-Фехнера. В 1760 г. Буггер
установил, что соотношение минимального воспринимаемого изменения освещенности
к величине освещенности есть величина постоянная в широких пределах
освещенности:
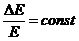
Такое же постоянство отношения минимально воспринимаемого прироста
раздражения ( ) к его исходной величине (R) было установлено Вебером и для
других сенсорных систем, то есть:
) к его исходной величине (R) было установлено Вебером и для
других сенсорных систем, то есть:
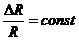
Так, например, прирост в весе груза, чтобы стать ощутимым, должен
превышать ранее действующий вес на 3%. Фехнер предположил, что минимальный
прирост ощущения (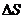 ) относительно исходного уровня ощущения (S) также постоянен,
тогда:
) относительно исходного уровня ощущения (S) также постоянен,
тогда:
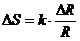 ,
,
где k- коэффициент пропорциональности.
Интегрируя это уравнение и проведя алгебраические преобразования, Фехнер
получил следующее выражение:
S = a lg R + b,
где a, b - постоянные.
Таким образом, согласно закону В-Ф, величина ощущения S пропорциональна
log силы раздражения. Эта зависимость обусловлена принципом кодирования
информации в рецепторном аппарате органов чувств.
Рецепторы представляют собой или специфические окончания афферентных
нервных волокон, или нервные окончания в соединении со специализированными
клетками, или структурами. Во всех случаях рецептор действует, как
преобразователь энергии раздражения в энергию нервного ответа.
При действии внешнего стимула изменяется ионная проницаемость рецепторных
мембран, что вызывает их деполяризацию. Это возникает под действием стимула и
деполяризация называется генераторным потенциалом (ГП). Величина ГП
пропорциональна log интенсивности действующего раздражителя. ГП рецептор клетки
генерирует в афферентном нервном волокне ПД, который служит сигналом,
передающим информацию в нервной клетке и нервной системе. Между величиной ГП и
частотой появления ПД в афферентном волокне, наблюдается линейная зависимость.
Нервное волокно может находиться в двух состояниях:
1 - в возбужденном, когда есть ПД;
2 - в невозбужденном, когда ПД отсутствует.
Таким образом, в нервной системе имеется дискретная двоичная форма
кодирования информации, как в цифровых ЭВМ. Как показывает эксперимент,
информация в нервной системе кодируется не последовательностью ПД, а их
частотой появления, как в аналоговых вычислительных машинах, где различные
модулированные показатели, представленные аналогами, изменяются пропорционально
величине модулированных факторов.
Поскольку между интенсивностью стимула и величиной ГП существует
логарифмическая зависимость, а ГП связан с частотой ПД линейно, интенсивность
стимула и частота ПД должны быть связаны логарифмической функцией. Работы Метьюса,
Грэма и других, выполненные с помощью микроэлектронной техники, подтверждают,
что частота пропорциональна логарифму величины действующего раздражителя.
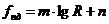 - частота ПД.
- частота ПД.
m,n=const.
Таким образом, по-видимому, данная зависимость преобразования
интенсивности действующего стимула (R) в частоту нервных импульсов в
афферентных волокнах и обуславливает логарифмическую зависимость величины
ощущения от интенсивности раздражителя.
54. Элементы биофизики слуха. Функции вспомогательных элементов наружного
и среднего уха
В органе слуха принято выделять три части:
·
наружное ухо;
·
среднее ухо;
·
внутреннее ухо.
Упрощенную схему органа слуха можно представить в виде:
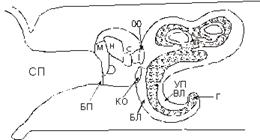
Схема органа слуха.
СП - слуховой проход; БП - барабанная перепонка; М - молоточек; Н -
наковальня;
С - стремечко; ОО - овальное окно; ВЛ - вестибулярная лестница;
УП - улитковый проток; БЛ - барабанная лестница; Г - геликотрема;
КО - круглое окно; НУ - наружное ухо; СУ - среднее ухо.
Слуховые рецепторы расположены во внутреннем ухе, тогда, как
вспомогательные элементы, способствующие слуховой рецепции, присутствуют во
всех частях органа слуха.
Наружное ухо включает ушную раковину и слуховой проход. С ним связано
свойство направленности слухового восприятия, под которым понимают лучшее
восприятие звуков, идущих в определенном направлении. Это обеспечивает
локализацию источника звука в пространстве (ототолику), кроме этого, слуховой
проход служит резонатором. Его резонансная частота у человека лежит в области
частот примерно 3 кГц, но резонансная кривая имеет уплощенную форму, поскольку
добротность СП, как колебательной системы, невелика. Разница в коэффициенте
передачи звуков резонансной и не резонансной частот не более 1 порядка.
Границей между наружным и средним ухом служит БП. НУ защищает БП от
механических повреждений и, поддерживая определенные микроклиматические условия
(температуру, влажность и пр.), стабилизирует ее механические свойства. Таким
образом, НУ выполняет следующие функции:
1. Локализация звука в пространстве.
2. Усиление звукового сигнала
(резонансное).
3. Механическая защита БП.
4. Обеспечение микроклимата, что
необходимо для нормального функционирования БП.
По СУ звук проводится цепочкой слуховых косточек: М, Н, С. рукоятка М
зафиксирована на БП, а его головка образует сустав с Н, который сочленяется с
С. Основание С погружено в ОО, которое служит входом во ВУ, поэтому система БП
и слуховых косточек СУ выполняет роль связующего звена между атмосферой и ВУ,
заполненным жидкостью. Если бы звук переходил из воздуха в жидкость
непосредственно, то интенсивность звуковой волны падала бы из-за сильного отражения
звука на границе раздела сред с различным акустическим импедансом [ ]. Жидкость обладает большей по сравнению с воздухом. БП
(исключает) и система слуховых косточек обеспечивает согласование воздуха и жидкости ВУ, вследствие
чего уменьшаются потери интенсивности звуковой волны при переходе из атмосферы
во ВУ.
]. Жидкость обладает большей по сравнению с воздухом. БП
(исключает) и система слуховых косточек обеспечивает согласование воздуха и жидкости ВУ, вследствие
чего уменьшаются потери интенсивности звуковой волны при переходе из атмосферы
во ВУ.
Согласование и усиление звука обеспечивается благодаря двум
обстоятельствам:
1. Поверхность БП примерно в 25 раз
больше площади ОО, за счет разницы в площадях на входе и выходе системы
механической передачи происходит усиление давления на жидкость ВУ, относительно
звукового давления, воздействующего на БП.
2. Слуховые косточки так расположены в
цепочке, что при их движении, они образуют рычаги силы и увеличивают давление
на основание С по сравнению с давлением на рукоятку М. Вся система передачи
звука в СУ работает наподобие гидравлического пресса с примерно 90 кратным
выигрышем в силе.
Максимальный коэффициент усиления давления характерен для звуковых волн с
f примерно 1 кГц. Звуковые колебания, как меньших, так и больших f, усиливаются
слабее, так как для них слуховые косточки СУ хуже согласовывают , и часть звуковых волн с f примерно
1 кГц, при их средней интенсивности, передается на ВУ практически без потерь.
Однако, и для них коэффициент передачи может снижаться и оказывается даже
меньше 1. Это имеет место в том случае, когда звук становится слишком
интенсивным. Это сопровождается ощущение покалывания, а затем и боли в ушах.
Понижение коэффициента передачи происходит при помощи тоненьких мышц, которые
прикрепляются к М и С. В ответ на сильные звуки, они рефлекторно сокращаются и
уменьшают подвижность системы слуховых косточек.
Итак, систему передачи звука, сосредоточенную в СУ, можно считать
механическим преобразователем (усилителем), обладающим переменными регулируемым
коэффициентом передачи давления с БП на жидкость ВУ. При разрушении слуховых
косточек слух полностью не теряется, но понижается примерно в 1000 раз.
СУ выполняет следующие функции:
1. Связующую между атмосферой и ВУ.
2. Согласования атмосферы (воздуха) и жидкости ВУ.
3. Усиление звукового сигнала.
4. Защитная, при больших интенсивностях
звука.
. Строение внутреннего уха
Внутреннее ухо (ВУ) человека размещается в пирамидальной части височной
кости. Часть этого причудливого лабиринта, содержащая слуховые рецепторы,
напоминает улитку в форме конусообразной спирали, образующей 2,5 витка.
Схематично костную часть улитки можно представить в виде:
Костная улитка.
а - стержень улитки; б - улитка в разрезе
Внутри улитки имеется просвет с круглым поперечным сечением, повторяющим
ее спиралевидное закручивание вокруг костного стержня. От стержня в просвет
улитки выступает костная спиральная пластинка (КСП), которая частично разделяет
просвет примерно на 1/3-1/2 на всем протяжении спиралевидного хода. Поперечный
разрез улитки имеет следующий вид:
Поперечный разрез улитки
- вестибулярная лестница, 2 - барабанная лестница, 3 - костная спиральная
пластинка, 4 - базилярная мембрана, 5 - мембрана Рейснера, 6 - покровная
пластинка (мембрана), 7 - улитковый проток, 8 - сосудистая полоска, 9 -
волосковые клетки внутреннего ряда, 10 - волосковые клетки наружного ряда, 11 -
нервные окончания, 12 - биополярные нейроны, 13 - аксоны слухового нерва
Между выступом костной спиральной пластинки (3) и противолежащей
поверхности улиткового хода натянута базилярная мембрана (4), которая
образована мягкими тканями БМ и дополняет костную спиральную пластинку, чем
достигается полное разделение просвета улитки. Кроме нее, от костной спиральной
пластинки отходит вестибулярная (преддверная) мембрана, которая называется
мембраной Рейснера (5). Эта мембрана образует острый угол с базилярной
мембраной, и она также направлена к наружной стенке улитки. Благодаря
существованию БМ и ВМ, спиралевидный ход внутри улитки разделен на три
параллельные канала, каждый из которых свернут в спираль. Один из них (нижний)
расположен между стенкой улитки и БМ. Он называется БЛ, другой канал - ВЛ (1)
заключен между верхней поверхностью улиткового хода и мембраной Рейснера. БЛ и
ВЛ сообщаются на верхушке улитки. Место перехода одной лестницы в другую
называют отверстием улитки или геликотремой (Г). Третий канал клиновидного
сечения называется улитковым протоком (7), он расположен между БМ и мембраной
Рейснера. Его третью стенку выстилает, так называемая, сосудистая полоска (8),
с функционированием которой связано поддержание определенного химического
состава жидкости - эндолимфы. Эндолимфа заполняет улитковый проток, и она по
своим свойствам существенно отличается от перелимфы, находящейся в БЛ и ВЛ.
Перелимфа сходна с межклеточной жидкостью, тогда как эндолимфа скорее
напоминает цитозоль (по содержанию , вязкости и т.д.).
В эндолимфу погружены специальные клетки слуховых рецепторов, которые
входят в состав Кортиева органа. Их опекальный полюс увенчан волосковоподобными
выростами плазмолеммы (цилии). В этой связи эти клетки называются волосковыми
клетками.
Наряду со слуховыми рецепторами в Кортиевом органе присутствуют
разнообразные опорные клетки, относящиеся к вспомогательным элементам ВУ.
Кортиев орган расположен на БМ, причем волосковые клетки ориентированы
перпендикулярно этой мембране. В каждой из двух улиток уха человека содержится
по 15500 волосковых клеток, из которых 3500 образуют внутренний ряд (9) и
12000- (3) наружных ряда (10). По форме волосковые клетки внутреннего ряда
подобны эллипсоиду (1), вертикальная ось которого составляет в среднем примерно
40 мкм, а горизонтальная примерно 10 мкм. Наружная волосковая клетка имеет
цилиндрическую форму (2).
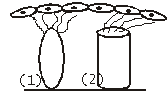
Волосковые клетки
- внутреннего ряда, 2 - наружного ряда
На базальном полюсе волосковой клетки сосредоточены синопсы с
терминальными веточками афферентных волокон, являющиеся периферическими
отростками нейронов, тела которых находятся в спиральном ганглии (12).
Центральные отростки этих биополярных нейронов формируют слуховой нерв (13), по
которому сигналы от слуховых рецепторов поступают в головной мозг. Каждая
волосковая клетка внутреннего ряда образует синопсы примерно с 10 нервными
окончаниями, но каждое из окончаний контактирует только с одной клеткой.
Афферентные волокна, иннервирующие волосковые клетки наружных рядов, сильно
ветвятся, и каждое из них имеет синоптические входы от многих таких клеток. В
этой связи, несмотря на обилие волосковых клеток в наружных рядах Кортиева
органа, основная часть (примерно 95%) афферентных волокон слухового нерва
млекопитающих отводит сигналы от волосковых клеток внутреннего ряда, которые и
преобразуют энергию механических колебаний.
. Распределение механических колебаний во внутреннем ухе
Под действием звука основание С колеблется, то вдвигаясь внутрь улитки,
то выходя из нее. Таким образом энергия механических колебаний передается
перелимфе ВЛ. Перелимфа может колебаться, благодаря двум обстоятельствам:
1. Жидкость, заполняющая ВЛ и БЛ,
составляет одно целое, поскольку лестницы сообщаются на верхушке улитки,
посредством Г.
2. Жесткий (костный) каркас, в который
заключена гидравлическая система внутреннего уха, имеет окно не только на входе
(ОО), но и на дальнем конце в БЛ (КО). КО затянуто мембраной. Оба окна
"смотрят" в полость СУ. Поскольку мембрана КО обладает упругостью
(эластичностью), начальное смещение перелимфы около ОО создает механическую
волну, распределяющуюся вдоль улиткового протока, который заполнен эндолимфой.
Среда, в которой распределяются механические колебания внутри улитки -
неоднородна. Она включает перелимфу, эндолимфу, разнообразные мембраны (БМ, ВМ
и покровную мембрану), которые сами по себе являются композиционными
материалами. Очень сложной биомеханической системой является БМ, с
расположенным на ней Кортиевым органом. Инерционность БМ увеличивается от
основания улитки к верхушке, тогда, как наиболее упругим ее участком является
начальный (около ОО), а по направлению к Г эластичность понижается на два
порядка.
Механические колебания внутри уха до настоящего времени не удалось
описать математически, однако, при световой микроскопии даже небольшой части
Кортиевого органа удалось наблюдать сложные колебания различных групп
волосковых клеток. В целом, колебательный процесс, возникающий во внутреннем
ухе под действием звуков, имеет характер бегущей волны. Волны такого типа
образуются в нелинейной механической системе, являясь результатом интерференции
различных колебаний, в которых приводятся различные элементы неоднородной
механической среды. Другими словами, одно и то же воздействие вызывает
неодинаковые колебания (с различными фазами и амплитудами, которые по-разному
направлены) того или иного элемента нелинейной механической системы, в
зависимости от его инерционных и упругих свойств. Вместе с тем, такие колебания
когерентны, и, накладываясь друг на друга, они интерферируют. В результате их
интерференции и образуется бегущая волна. В отличие от волновых процессов в
однородной среде, бегущей волне не всегда свойственно максимальное смещение в
начальном участке ее распределения и последующее угасание с определенным
декрементом. Бегущая волна достигает своей максимальной амплитуды на различном
удалении от входа в нелинейную механическую систему, а именно, в той области,
где образуется интерфереционный максимум, при суперпозиции различный колебаний,
которые возникают в неоднородной среде. Необходимо отметить, что местоположение
максимального смещения в композиционной среде, при периферическом воздействии
на нее механических сил, зависит от частоты сигнала.
Бегущая волна, возникающая в БМ под действием звуков НЧ, достигает
максимального значения (амплитуды) ближе к ОО, тогда как звуки НЧ вызывают
максимальное смещение тех участков БМ, которые находятся около Г. Звуковые тоны
f = 1,6 кГц максимально смещают середину БМ человека. Таким образом, БМ, как
композиционная среда с нелинейными механическими свойствами, функционирует
наподобие системы фильтров, в которых расположение максимального отклика
зависит от частоты звукового тона.
. Слуховая рецепция
Благодаря зависимости, местоположение максимальной амплитуды бегущей
волны на БМ от частоты вызывающей ее звукового тона, происходит распределение
этих частот между различными участками Кортиевого органа. Его волосковые клетки
возбуждаются, главным образом, в местах максимального смещения БМ,
следовательно, звуковой тон каждой частоты возбуждает соответствующие слуховые
рецепторы. В этой связи в У осуществляется первый этап частотного анализа
звука, который основывается на пространственном разграничении участков БМ,
колеблющихся с неодинаковой амплитудой, под действием звукового тона
определенной частоты. Звуки пороговой интенсивности, вызывающие бегущие волны,
амплитуда которых в Кортиевом органе не превышает  м. При усилении звуков, амплитуда
колебаний БМ больше этой величины всего на 1-2 порядка, следовательно, слуховой
рецептор реагирует на ничтожные механические перемещения. Начало этой реакции
связано с движением волосков (цилий) на опекальном полюсе волосковой клетки эти
смещения обусловлены распределением бегущей волны по БМ. Среди цилий в каждой
внутриволосковой клетке различают одну длинную (киноцилия) и множество
(примерно 100-120) коротких (стереоцилий). Рецепция звука обусловлена
воздействием механических усилий на стереоцилии.
м. При усилении звуков, амплитуда
колебаний БМ больше этой величины всего на 1-2 порядка, следовательно, слуховой
рецептор реагирует на ничтожные механические перемещения. Начало этой реакции
связано с движением волосков (цилий) на опекальном полюсе волосковой клетки эти
смещения обусловлены распределением бегущей волны по БМ. Среди цилий в каждой
внутриволосковой клетке различают одну длинную (киноцилия) и множество
(примерно 100-120) коротких (стереоцилий). Рецепция звука обусловлена
воздействием механических усилий на стереоцилии.
Стереоцилии прикасаются к покровной мембране, которая имеет желеобразное
состояние. Один край покровной мембраны закреплен на костной спиральной
пластинке, а другой, свободный, нависает над свободными клетками. Модель изгиба
стереоцилий под действием звуковых тонов, схематически можно представить в виде:
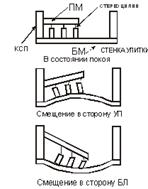
Модель изгиба стереоцилий под действием звука
КСП - костная спиральная пластинка; БМ - базилярная мембрана; ПМ -
покровная мембрана; СУ - стенка улитки, к которой прикрепляется базилярная
мембрана; С - стереоцилии волосковой клетки
При совместных колебаниях БМ и ПМ происходит изгиб стереоцилий. Небольшое
смещение их свободных концов приводит к значительной конформационной
перестройке молекул плазмолеммы волосковой клетки в тех местах, где залегают
"корни" изогнувшихся волосков, поскольку стереоцилиям присущи
свойства микроочагов.
Конформации мембраных макромолекул обусловливают изменение проницаемостей
для ионов и, следовательно, приводят к возникновению ионного тока через
мембрану волосковой клетки. Это сопровождается сдвигом разности потенциалов на
мембране, когда стереоцилии изгибаются в сторону киноцилий, рецепторная клетка
деполяризуется. При их противоположном смещении наблюдается гиперполяризация.
Только деполяризация волосковой клетки обеспечивает усиление импульсации в
слуховом нерве, поэтому, именно деполяризационный сдвиг мембранного потенциала
называется рецепторным потенциалом волосковой клетки - РП.
Уровень ПП волосковой клетки относительно межклеточной среды (перелимфы),
находится в пределах от -50 до -70 мВ. Значение рецепторного потенциала зависит
от интенсивности звука, и его величина составляет примерно 10 мВ. Максимальная
величина РП, зарегистрированного с помощью микроэлектронной техники, составляет
примерно 24 мВ. РП электротонически распределяется по плазмолемме волосковой
клетки от ее опекального полюса к базальному. Там деполяризация приводит к
высвобождению медиатора (определенное химическое вещество), поступающего далее
через синоптическую щель на субсиноптическую мембрану, которая принадлежит
чувствительному нервному окончанию, контактирующему с данной волосковой
клеткой. Под действием медиатора, на субсиноптической мембране возникает
генераторный потенциал - ГП. Этот ГП электротонически распространяется на
внесиноптические участки афферентных волокон, где вызывает образование ПД. В
таком виде нервная импульсация направляется в ЦНС по слуховому нерву.
Следовательно, по механизму преобразования адекватного стимула (звука) в
нервные импульсы, слуховой рецептор является типичным представителем вторичночувствующих
рецепторов.
Каждая волосковая клетка соединена с нервными окончаниями, при этом,
каждое волокно слухового нерва начинается от узкого ограниченного участка
Кортиевого органа, в отдельных случаях, от одной волосковой клетки. Поскольку
слуховые рецепторы расположены в том или ином месте БМ возбуждается звуками
определенных частот, то каждая небольшая группа нервных волокон слухового нерва
проводит импульсы в ответ на звуки преимущественно одной частоты. Эта частота
называется характерической частотой волокна. Если звук представляет собой
сложные колебания, то в слуховом нерве активизируются все волокна,
характерические частоты которых соответствуют гармоническому спектру сложного
звука. Следовательно, на уровне слуховых рецепторов звуки разлагаются в
гармонический спектр. Длительность звукового сигнала кодируется временем
активации афферентных волокон, которые входят в состав слухового нерва.
. Роль эндокохлиарного потенциала (ЭКП) в слуховой рецепции
Между эндолимфой и перелимфой в У поддерживается разность потенциалов в
пределах от 60 до 80 мВ. Эту разность называют ЭКП. Эндолимфа несет
положительный потенциал относительно перелимфы. Источником ЭКП служит
сосудистая полоска, выстилающая наружную стенку улиткового протока (8). Падение
ЭКП наблюдается при нарушении кислородного снабжения сосудистой полоски, или
при нарушении этой полоски (механическое повреждение).
Клетки, входящие в состав сосудистой полоски, содержат очень крупные
митохондрии. Эта структурная особенность сосудистой полоски в совокупности с
данными о роли ее в клеточном дыхании для поддержания разности потенциалов
между эндолимфой и перелимфой, позволяют высказать предположение, что ЭКП по
своей природе может быть аналогичен протонному потенциалу на митохондриальной
мембране.
Звуки вызывают колебания ЭКП, амплитуда которых не превосходит 1% его
постоянного уровня, то есть, эти колебания составляют 0,6-0,8 мВ. Это явление
получило название микрофонного эффекта улитки, а сами колебания ЭКП принято
называть микрофонными потенциалами. Микрофонный эффект У положен в основу
телефонной теории слуха, согласно которой, частотный анализ звуков связан не с
механическими колебаниями структур ВУ, а с возбуждением слуховых рецепторов
микрофонными потенциалами. Частота этих потенциалов совпадает с гармоническим
спектром звуковых тонов. В настоящее время ЭКП рассматривается, как важное
условие нормального функционирования слуховых рецепторов, но не ведущий ее
механизм.
Благодаря существованию ЭКП, поддерживается очень большая разность
потенциалов (до 150 мВ) между эндолимфой и цитоплазмой волосковой клетки. Эта
разность образуется в результате суммирования ПП волосковой клетки (разности
потенциалов между ее цитоплазмой и перелимфой) и ЭКП.
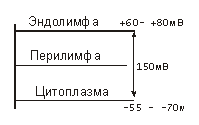
Уровень электрических потенциалов волосковой клетки
По существу, разность потенциалов между эндолимфой и цитоплазмой
волосковой клетки выполняет роль ее ПП, относительно которой генерируется
рецепторный потенциал. Столь значительный ПП не имеет ни одна другая клетка в
организме человека. Это высокая крутизна переднего фронта ЭКП делает
плазмолемму волосковой клетки чрезвычайно нестабильной и способной давать
резкие и быстрые сдвиги мембранного потенциала, при малых изменениях
электрического сопротивления мембраны, которые происходят в ответ на
незначительные смещения стереоцилий.
59. Основы физиологической акустики
Физической акустике принято сопоставлять объективные (физические) и
субъективные (психологические) характеристики звука.
|
Физические характеристики
звука
|
Психологические
характеристики звука
|
|
1. Гармонический спектр
(характеризуется суммой всех частот 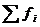 ): а)
основная гармоника; б) высшие гармоники. ): а)
основная гармоника; б) высшие гармоники.
|
1. Акустический спектр,
характеризуется: а) основной тон (высота); измеряется в [мел] или [барк]; б)
обертоны (тембр).
|
|
2. Интенсивность (плотность
потока звуковой энергии), обозначается  . .
|
2. Громкость, измеряется в
[сон].
|
Звук, при распределении в среде представляет собой волны сгущения и
разряжения ее частиц. В участках сгущения давление больше, чем в участках
разряжения. Амплитуда периодических колебаний давления в среде, происходящих
под действием распространяющего звука, называют звуковым давлением - р. Оно
имеет размерность [Па] или [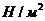 ] Звуковым давлением определяется интенсивность звука,
которая определяется так:
] Звуковым давлением определяется интенсивность звука,
которая определяется так:
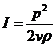
- давление;- скорость распределения звука в данной среде;
- плотность среды [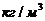 ].
].
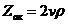 - акустический импеданс среды.
- акустический импеданс среды.
Кроме того, I, как вектор Умова, можно рассчитать по формуле:
 , если известна w - объемная плотность энергии звука, то
есть, энергия, приходящаяся на единицу объема среды, в которой звук
распространяется. Для установления взаимосвязи между интенсивностью и
громкостью, вводятся еще две физические величины, которые характеризуют звук:
, если известна w - объемная плотность энергии звука, то
есть, энергия, приходящаяся на единицу объема среды, в которой звук
распространяется. Для установления взаимосвязи между интенсивностью и
громкостью, вводятся еще две физические величины, которые характеризуют звук:
1. Уровень интенсивности - L.
2. Уровень громкости - Е.
L, или уровень звукового давления, определяется по формуле:
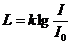
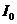 - эталонная интенсивность;
- эталонная интенсивность;
- любой исследуемый звук.
обычно выбирают меньше  Вт/
Вт/ . Такая интенсивность чистого тона, f
= 1 кГц, установлена, как средний порог слуховой чувствительности для человека.
Эта интенсивность соответствует звуковому давлению, равному
. Такая интенсивность чистого тона, f
= 1 кГц, установлена, как средний порог слуховой чувствительности для человека.
Эта интенсивность соответствует звуковому давлению, равному  Па. При к = 1, единицей уровня
интенсивности L служит [Б] бел. При к = 10, единицей уровня интенсивности L
служит [дБ], децибел. В медицине принято использовать децибельную шкалу. Если
Па. При к = 1, единицей уровня
интенсивности L служит [Б] бел. При к = 10, единицей уровня интенсивности L
служит [дБ], децибел. В медицине принято использовать децибельную шкалу. Если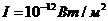 , то L = 0, следовательно, 3 величины
, то L = 0, следовательно, 3 величины
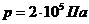 и , и L=0 дБ - характеризуют один и тот
же звук. При
и , и L=0 дБ - характеризуют один и тот
же звук. При 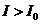 , L принимает положительно значение, при
, L принимает положительно значение, при 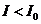 , L < 0. Чем меньше L, тем лучше
(выше) слуховая чувствительность. Человек лучше всего слышит звуковые тоны с f
от 3 до 4 кГц.
, L < 0. Чем меньше L, тем лучше
(выше) слуховая чувствительность. Человек лучше всего слышит звуковые тоны с f
от 3 до 4 кГц.
Уровень громкости (Е) - это выражение различий в восприятии человеком
звуков разной частоты. Уровнем громкости звука данной f называют уровень
интенсивности звукового тона с f = 1 кГц, громкость которого при сравнении на
слух эквивалентна громкости исследуемого звука.
Размерность физической величины Е - фон, - для звуков, частотой f = 1
кГц, значение Е и L совпадают. Так, если первая из них L составляет 0 дБ, то Е
= 0 фонов (на f = 1 кГц). Средний порог слухового восприятия у человека
составляет примерно 4 фонов. Болевые ощущения в ухе возникают под действием
звуков, уровень громкости которых достигает примерно 130 фонов. Таким образом,
все, что слышит человек, ограничивается звуковыми тонами, частота которых
находится в диапазоне от 16 Гц до 20 кГц, а Е - от 4 до 130 фонов.
Громкость звука является психологической (субъективной) характеристикой.
Она оценивается по шкале сонов. Для выражения громкости того или иного звука в
сонах, за эталон принимается звуковой тон с f = 1 кГц, при уровне интенсивности
L = 40 дБ. Дав испытуемому послушать эталонный тон, врач предъявляет ему
тестированный звук, при этом, он просит определить, во сколько раз тестируемый
звук громче или тише эталонного. Так, например, громкость тестируемого звука,
если он слышит в 2 раза громче эталонного тона, будет равен 2 сонам, а, если в
2 раза тише - то 0,5 сон.
. Вестибулярная сенсорная система
Вестибулярный анализатор участвует в регуляции положения и движения тела
в пространстве, в поддержании равновесия и регуляции мышечного тонуса.
Периферический отдел анализатора представлен рецепторами, расположенными
в преддверии улитки, находящейся внутри височной кости. На рис. 11 изображено
взаимное расположение улитки и ее преддверия.
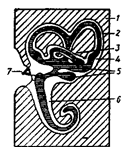
Рис. 11. Улитка и ее преддверие: 1 - височная кость; 2
- вертикальные полукружные каналы; 3 - горизонтальный полукружный канал; 4 -
купула; 5 - макулы вестибулярного аппарата; 6 - улитка; 7 - полость среднего
уха
Вестибулярные рецепторы возбуждаются при изменении скорости вращательного
движения, прямолинейном ускорении, изменении направления силы тяжести,
вибрации. Нервные импульсы, возникшие в этих рецепторах, поступают по вестибулярному
нерву в ствол мозга, а оттуда по проводящим путям - к центральному отделу
анализатора, расположенному в передних отделах височной доли коры головного
мозга. В результате возбуждения нейронов этого отдела коры возникают ощущения,
дающие представления о положении тела и отдельных его частей в пространстве,
способствующие сохранению равновесия и поддержанию определенной позы тела в
покое и при движении.
При этом действуют особые механизмы трансформации внешней силы в
механическое воздействие на так называемые волоски рецепторной клетки,
выступающие из ее апикального полюса в эндолимфу.
Поэтому рецепторные клетки называются волосковыми.
Будучи расположенными в полукружных каналах, они реагируют на угловые
перемещения эндолимфы, а значит, и всего тела человека и животного. В
преддверии улитки, кроме полукружных каналов, находятся еще две структуры:
маточка и мешочек.
Их волосковые клетки реагируют на прямолинейные перемещения тела.
Рецепторным органом маточки и мешочка (рис. 12, а) являются так называемые
пятна.
Волоски клеток, образующих пятна, направлены внутрь заполненной
эндолимфой полости, где они погружены в желеобразную массу, заключающую в себе
кристаллики кальциевых солей (камешки - отолиты). Вследствие своеобразия этих
кристалликов пятна нередко называют отолитовым прибором.
При наклоне головы под действием силы земного притяжения происходит
смещение отолитов, что ведет к сгибанию волосков.
Сгибание волосков, как у других механорецепторных клеток, передает сигнал
на контактирующие с рецепторной клеткой нервные окончания. Регистрация
активности последних показала, что ритмическая импульсная активность
генерируется рецептором и в том случае, когда волоски выпрямлены. При сгибании
их в одну сторону она прекращается, а при сгибании в другую - учащается.
Так как макулы мешочка и маточки расположены в разных плоскостях - одна
примерно в горизонтальной, а другая - в вертикальной, то при любом положении
головы в пространстве будет иметь место особое сочетание импульсной активности
в иннервирующих их афферентных нервных волокнах, передающих в нервные центры
информацию об этом положении.
Три полукружных канала, расположенные во взаимно перпендикулярных
плоскостях, образуют декартову систему координат.
Их механорецепторными органами являются так называемые гребешки,
образованные волосками рецепторных клеток и желеобразной массой (рис. 12, б,
в).
Рис. 12. Упрощенная модель вестибулярного аппарата: а
- аппарат, воспринимающий линейные ускорения (1 - отолиты, 2 - отолитовая
мембрана, 3 - волосковые клетки); б, в - аппарат, воспринимающий угловые
ускорения (1 - ампула, 2 - купула, 3 - полукружный канал с перилимфой, 4 -
волосковая клетка, 5 - гребешок); б - в покое, в - при угловом перемещении
Гребешки сосредоточены в расширениях (ампулах) каждого из трех каналов.
Погруженные в желеобразную массу, гребешки перегораживают канал подвижной
желеобразной перегородкой (купулой). Как показали прямые наблюдения, смещение
купулы и, следовательно, сгибание или разгибание волосков происходит при
перемещениях эндолимфы в канале. В свою очередь, такие перемещения возникают в
случае ускорения или замедления вращения головы в плоскости, в которой
расположен соответствующий канал (вследствие инерции эндолимфы). При переходе
вращения в равномерное купула медленно возвращается в свое исходное положение.
Таким образом, эта рецепторная система оказывается сенсором вращательного
ускорения. Она сигнализирует о нем разрядом импульсов, частота которого
уменьшается и приобретает стационарный характер при переходе к равномерному
вращению или к покою. В физиологических условиях ускорения вращения являются
постоянно встречающимися раздражителями, возникающими при каждом движении
головы. Равномерное же вращение - состояние, как правило, искусственное, возникающее
в связи с развитием техники, поэтому оно и не является адекватным раздражителем
для сенсорных клеток полукружных каналов.
61. Элементы биофизики зрения. Светопреломляющая система глаза
Органом зрения принято называть глаз. Однако, в зрительную сенсорную
систему (зрительный анализатор) вместе с фоторецепторами (палочками и
колбочками) и рядом нейронов, которые находятся в сетчатой оболочке глаза
входят определенные отделы ЦНС. В то же время, глаз включает не только
фоторецепторы и нейроны, но также разнообразные вспомогательные элементы
зрительной рецепции. Эти вспомогательные элементы сами не воспринимают свет и
не участвуют в переработке (передаче) сигналов от фоторецепторов, но
способствуют восприятию света. Важнейшим вспомогательным аппаратом фоторецепции
является диоптрическая светопреломляющая система глаза. Упрощенная схема
разреза глаза человека имеет вид:
Схема горизонтального разреза глаза человека
- роговица; 2 - хрусталик; 3 - стекловидное тело, камерная влага;
- сосудистая оболочка; 8 - склера; OO’ - главная оптическая ось;’
-зрительная ось; 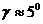 - угол между OO’ и KK’.
- угол между OO’ и KK’.
Светопреломляющая система глаза фокусирует изображение окружающего мира
на сетчатой оболочке (сетчатке), в которой расположены фоторецепторы.
Диоптрический аппарат глаза подобен сильной собирающей линзе, и, при
рассмотрении далеких предметов, сводит в точку на сетчатке входящие в глаз параллельные
лучи, то есть, действует наподобие объектива зрительной трубы. Фокусированные
изображения осуществляются за счет преломляющей системы глаза, к которой
относятся:
·
роговица;
·
хрусталик;
·
камерная плата;
·
стекловидное
тело.
Преломляя световые лучи, они обеспечивают формирование на сетчатке
действительного уменьшенного и обратного (перевернутого) изображения
рассматриваемого предмета. Отрезок прямой, проходящий через геометрические
центры роговицы и хрусталика, называют главной оптической осью глаза (ОО’). Она
объединяет весь его диоптрический аппарат в центрированную светопреломляющую
систему. Центрированной называют такую систему светопреломляющих поверхностей,
геометрические центры которых лежат на одной прямой. Обратимся к идеальной
центрированной системе Гаусса и попытаемся применить ее к человеческому глазу.
Центрированная диоптрическая система при построении изображения предметов
рассматривается, как одно целое, и ход лучей в ней определяется при помощи
шести, так называемых кардинальных точек, которые лежат на оси ОО’. К этим
точкам относятся два главных фокуса (передний F1 и задний F2), две главные
точки (Н1 и Н2) и две узловые точки (N1 и N2).
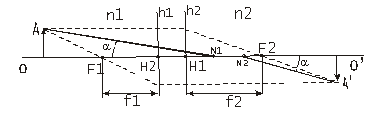
Схема построения оптического изображения
Главными фокуcами называют точки (F1 и F2), в которых после преломления
собираются лучи, входящие в систему параллелей, так как коэффициенты
преломления сред (n), находящихся перед глазом 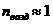 , и внутри глаза (n=1,336), не
одинаковы, то и фокусные расстояния
, и внутри глаза (n=1,336), не
одинаковы, то и фокусные расстояния 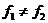 . Размеры изображений предметов,
построенных на h1 и h2, одинаковы. В местах пересечения главных плоскостей с
главной осью расположены главные точки Н1 и Н2, от этих точек отсчитываются
фокусные расстояния: от Н1 - переднее, f1, от Н2 - заднее f2.
. Размеры изображений предметов,
построенных на h1 и h2, одинаковы. В местах пересечения главных плоскостей с
главной осью расположены главные точки Н1 и Н2, от этих точек отсчитываются
фокусные расстояния: от Н1 - переднее, f1, от Н2 - заднее f2.
Среди лучей, исходящих от предмета, всегда находится такой, который после
преломления диоптрической системы, покидает ее без изменения первоначального
направления. Этот луч претерпевает в системе только смещение, вследствие чего и
распространяется по входу и выходу из нее, параллельно исходному направлению.
Точки, в которых продолжение падающего луча и выходящего луча пересекают
главную ось, называются узловыми точками. N1 - передняя узловая точка, N2 -
задняя. Через эти точки падающий и преломленный лучи проходят под одинаковым
углом по отношению к главной оси (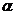 ), применительно к глазу, он
называется углом зрения или зрительным углом.
), применительно к глазу, он
называется углом зрения или зрительным углом.
. Тонкая и толстая линзы
Если вся центрированная преломляющая система находится в однородной среде
(n1 = n2), то N1 совпадает с Н1, а N2 с Н2. Такой относительно простой
центрированной диоптрической системой является тонкая линза, когда обе ее
поверхности контактируют с воздухом. В тонкой линзе можно пренебречь ее
толщиной по сравнению с расстоянием соответствующих фокусов от передней и
задней поверхности, тогда, в этом случае, в центрированной системе точки N1 и
N2, Н1 и Н2 являются центром оптической линзы, то есть сливаются в одну. При
построении изображений, которые получаются в тонкой линзе, окруженной
однородной средой, расстояние от оптического центра до предмета и расстояние до
изображения от оптического центра, связаны с фокусным расстоянием по следующей
формуле:
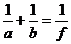 - формула тонкой линзы.
- формула тонкой линзы.
 - расстояние от предмета до оптического центра;- расстояние
от оптического центра до изображения.
- расстояние от предмета до оптического центра;- расстояние
от оптического центра до изображения.
Для толстой линзы, чему можно уподобить центрированную систему глаза,
формулы, связывающие и b, гораздо сложнее, они включают все кардинальные точки
центрированной диоптрической системы. Для упрощения расчетов прибегают к
моделям реальных диоптрических систем. При этом стремятся обойтись в модели
минимальным количеством кардинальных точек.
. Приведенный редуцированный глаз человека
Моделью светопреломляющего аппарата органа зрения служит, так называемый,
приведенный редуцированный глаз. Редукция означает упрощение, то есть сведение
сложного к простому, более доступному для анализа. Физическим аналогом
приведенного редуцированного глаза является стеклянная линза, которая одной
поверхностью контактирует с воздухом, а другой - с жидкостью, обладающей n =
1,336. Первая поверхность направлена в пространство предметов, а другая
поверхность линзы - к пространству изображений.
Диоптрический аппарат глаза не поддается точному математическому
описанию. Это связано с тем, что глаза разных людей сильно различаются, а также
из-за подвижности хрусталика, и по ряду других причин. Так, например, главная
оптическая ось глаза человека проводится довольно приблизительно. Она не
совпадает со зрительной осью КК’, которая проходит через геометрические центры
роговицы и желтого пятна сетчатки. В направлении зрительной оси глаз имеет
наилучшую разрешающую способность. Угол между главной оптической осью ОО’ и
KK’, как правило не превышает 5 градусов. Он обозначается  обычно учитывается в офтальмологии
при назначении очков.
обычно учитывается в офтальмологии
при назначении очков.
Итак, человеческий глаз можно рассматривать в качестве центрированной
преломляющей системы весьма условно. На рисунке приведена количественная
характеристика одной из наиболее адекватных физических моделей человеческого
глаза.
Глаз как центрированная светопреломляющая система
Из рисунка видно, что расстояние между Н1 и Н2 составляет примерно 0,25
мм. Столь близкое расстояние допускает замену двух главных плоскостей (h1 и h2)
на одну главную плоскость. Точки N1 и N2 практически также совпадают в единую
узловую точку глаза, ее называют оптическим центром глаза (между N1 и N2). В
замене каждой из названных пар кардинальных точек на единые угловую и узловую и
состоит смысл редукции центрированной диоптрической системы реального глаза. В
схеме Гульстранда единая плоскость h и единая узловая точка N находятся на
расстоянии 1,6 мм и 7,5 мм соответственно от передней поверхности роговицы. Все
расстояния, кроме фокусных (f1, f2), отсчитываются от передней поверхности
роговицы. По ходу лучей внутрь глаза эти расстояния имеют знак "+". В
противоположном (наружном) направлении расстояния имеют знак "-".
Числа, характеризующие расстояние единых главной и узловой точек глаза,
необходимы для расчета диоптрического аппарата человеческого глаза.
. Биофизический механизм восприятия света рецепторами
Свет, сфокусированный диоптрическим аппаратом глаза на сетчатке,
возбуждает ее фоторецепторы. Он достигает фоторецепторов, пройдя всю толщу
сетчатки, поскольку палочки и колбочки располагаются в тех слоях сетчатки, которые
прилежат ближе к сосудистой оболочке. Свет проходит в сетчатке следующие слои,
прежде чем достигнуть фоторецепторов:
1. Слой ганглеозных клеток.
2. Внутренний синоптический слой.
3. Внутренний ядерный слой, состоящий из
амакриловых клеток, горизонтальных клеток (Г), биполярных нейронов (Б).
4. Наружный синоптический слой палочек и
колбочек (П и К).
5. Внутренний ядерный слой П и К.
Каждый глаз человека состоит из примерно 125 млн палочек и примерно 5 млн
колбочек.
Фоторецепторы П или К схематично можно представить в виде:
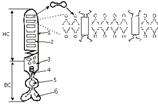
Схема строения фоторецепторной клетки (палочки или колбочки)
- наружная мембрана П или К; 2 - диски; 3 - митохондрии; 4 - аппарат
Гольджи; 5 - ядро клетки; 6 - синоптическая зона: НС - наружный сегмент; ВС -
внутренний сегмент.
Диаметр П (К) составляет примерно 2 мкм, длина - примерно 50-60 мкм.
Длина НС составляет примерно 20-30 мкм. Вся цитоплазма НС заполнена
своеобразными органеллами, которые называют дисками. Промежутки между ними не
превышают 15 нкм. В НС каждой П (К) содержится от 700 до 1000 дисков, которые
напоминают собой сплющенные воздушные шары, сложенные в стопку. Диаметр каждого
диска почти соответствует диаметру наружного слоя (примерно 2 мкм), а толщина
диска составляет примерно 15-18 нм. Его внутреннее пространство представляет
собой узкую щель, шириной = 1 нм. Эта щель расширяется до нескольких нм у краев
диска. Таким образом, на внутридисковое содержимое вместе с цитозолем
приходится не более половины объема НС, а другую половину занимают мембраны
дисков.
НС буквально нафарширован мембранами, и именно в них осуществляется
первичный процесс восприятия света. В этой связи, мембрану диска называют
фоторецепторной мембраной.
Молекулярная организация фоторецепторной мембраны у позвоночных и
беспозвоночных животных практически одинакова. Подобно всем БМ, она состоит из
липидов и белков (протеинов), однако, ей присущи две кардинальные отличительные
особенности. Во-первых, ее основным белковым компонентом является зрительный
пигмент - хромогликопротеид родопсин. Его концентрация достигает 90% от общего
содержания протеинов в фоторецепторной мембране. Во-вторых, более 40%
жирокислотного состава фосфолипидов, образующих дисковые мембраны, приходится
на полиненасыщенную декозагексоеновую кислоту, в состав которой входит 22 атома
углерода с присутствием 6 двойных связей. Кроме нее, в состав мембранных
фосфолипидов входят и другие полиненасыщенные длинноцепочечные жирные кислоты,
содержащие от 4 до 6 двойных связей. Вместе они составляют более половины всех
жирных кислотных остатков. Высокая степень ненасыщенности жирнокислотных
"хвостов" фосфолипидных молекул придает гидрофобному бимолекулярному
каркасу фоторецепторной мембраны необычайно высокую подвижность, вследствие
чего данная мембрана обладает малой вязкостью по сравнению с другими
мембранами.
. Реакция родопсина на действие света
Первичный механизм возбуждения П светом связан со сложными превращениями
родопсина в фоторецепторной мембране. Родопсин - это высокомолекулярное
соединение, состоящее из двух основных компонентов:
1. Альдегид витамина А (ретиналь).
2. Липопротеин под названием опсин
(белок глаза).
Спектр поглощения родопсина имеет вид:
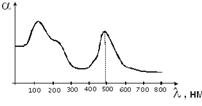
Спектр поглощения родопсина
Спектр поглощения родопсина хорошо приспособлен к восприятию света в
достаточно широкой области солнечного излучения, но лучше всего этот зрительный
пигмент поглощает свет в зеленой части этого спектра (500 нм). Красные и синие
лучи он отражает, причем, в отраженном свете преобладает красный компонент,
хотя присутствует и синий. Поэтому пигмент имеет пурпурный цвет, определивший и
его название. Родопсин относится к наиболее интенсивно окрашиваемым соединениям
из всех известных в настоящее время в органической химии. Возможность
поглощения фотонов родопсина близка к теоретически возможному значению.
В темноте ретиналь пребывает в 11 цис-форме, для которой характерна
некоторая скрученность молекулы. Скрученность объясняется тем, что в планарной
форме между близко расположенными метильной группой у углерода (С13) и
водородом у С10, возникает сильное отталкивание. Сила отталкивания не допускает
плоского расположения -связи. Скрученность в молекуле 11 цис-ретиналя обеспечивает
наибольшую энергию взаимодействия с опсином, вследствие чего их комплекс
(ретиналя и опсина) весьма устойчив. Для его разрушения необходимо
преобразовать 11 цис-ретиналь в другой изомер, в так называемый полностью
транс-ретиналь. Такое преобразование из цис- в транс-ретиналь происходит под
действием света. Этот процесс называется фотоизомеризацией.
При поглощении фотона (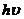 ), молекула родопсина претерпевает электрический переход
"
), молекула родопсина претерпевает электрический переход
"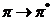 ", при этом, молекула родопсина
оказывается в синглетном возбужденном состоянии. Необходимо отметить, что в
органических молекулах -орбитали, которые свойственны основному энергетическому
состоянию, являются связывающими, то есть, они стремятся к перекрытию и
образованию второй связи. Напротив, -орбитали, характерные для
возбужденного состояния молекулы, относятся к разрыхляющим орбиталям. Их
перекрытие энергетически невыгодно. В силу чего, части молекулы, соединенные
двойной связью с участием -орбитали, разворачиваются на 90 градусов, и двойная связь
превращается в одинарную. В этой связи, в молекуле ретиналя, возбужденной
светом, происходит поворот атомов вокруг двойной связи. Такое измененние
пространственного расположения атомов, входящих в состав ретиналя, требует
затрат энергии. Эти энергозатраты характеризуются преодолением потенциального
барьера. Для разворота атома вокруг двойной связи в 11 цис-ретинале достаточно
энергии одного фотона. Для фотоизомеризации достаточно и времени, в течение
которого молекула родопсина пребывает в сигнетном возбужденном состоянии (
", при этом, молекула родопсина
оказывается в синглетном возбужденном состоянии. Необходимо отметить, что в
органических молекулах -орбитали, которые свойственны основному энергетическому
состоянию, являются связывающими, то есть, они стремятся к перекрытию и
образованию второй связи. Напротив, -орбитали, характерные для
возбужденного состояния молекулы, относятся к разрыхляющим орбиталям. Их
перекрытие энергетически невыгодно. В силу чего, части молекулы, соединенные
двойной связью с участием -орбитали, разворачиваются на 90 градусов, и двойная связь
превращается в одинарную. В этой связи, в молекуле ретиналя, возбужденной
светом, происходит поворот атомов вокруг двойной связи. Такое измененние
пространственного расположения атомов, входящих в состав ретиналя, требует
затрат энергии. Эти энергозатраты характеризуются преодолением потенциального
барьера. Для разворота атома вокруг двойной связи в 11 цис-ретинале достаточно
энергии одного фотона. Для фотоизомеризации достаточно и времени, в течение
которого молекула родопсина пребывает в сигнетном возбужденном состоянии (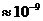 с), поскольку цис-тран-изомеризация
ретиналя происходит всего за 0,1-0,3 пс, квантовый выход фотоизомеризации 11
цис-ретиналя в родопсине достигает 0,7.
с), поскольку цис-тран-изомеризация
ретиналя происходит всего за 0,1-0,3 пс, квантовый выход фотоизомеризации 11
цис-ретиналя в родопсине достигает 0,7.
Фотоизомеризация - есть первая и единственная фотохимическая реакция в
зрительном акте. Она протекает внутри отдельной молекулы родопсина и, в отличие
от фотосинтеза, не сопровождается никакими химическими реакциями. Зрение
отличается от фотосинтеза, который происходит в растениях, еще и тем, что между
молекулами родопсина невозможен перенос энергии, так как, в фоторецепторной
мембране молекулы родопсина удалены друг от друга на расстояние более 7 нм.
Непосредственным следствием фотоизомеризации, приводящей к появлению у человека
ощущения света, служат конформационные перестройки молекулы родопсина. Дело в
том, что пространственно 11 цис-ретиналь точно соответствует конфигурации
активного центра. При образовании транс-ретиналя, это соответствие
(комплементарность) нарушается. В результате чего ретиналь отщепляется от
опсина, и родопсин обесцвечивается, то есть, претерпевает фотолиз.
Вне действия света, распавшийся родопсин восстанавливается. В ходе
восстановления, транс-ретиналь преобразуется в цис-форму. Такой переход
обратной фотоизомеризации называется реизомеризацией. У позвоночных животных
транс-ретиналь, отщепившись от опсина, поступает в пигментный эпителий
сетчатки, где подвергается ферментативным превращениям. Из пигментного эпителия
фоторецептором (П, К) доставляется 11 цис-ретиналь. Каждая палочка ежесекундно
синтезирует 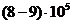 молекул родопсина из поступающего в нее 11 цис-ретиналя,
который соединяется с опсином. Суточный синтез этого зрительного пигмента в
целом оценивается величиной примерно
молекул родопсина из поступающего в нее 11 цис-ретиналя,
который соединяется с опсином. Суточный синтез этого зрительного пигмента в
целом оценивается величиной примерно 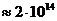 молекул.
молекул.
Конформационная перестройка родопсина, называющаяся фотоизомеризацией
ретиналя, довольно существенна. Естественно, что перестройку сопровождают
значительные изменения физико-химических свойств фоторецепторной мембраны, в
которую встроен родопсин. Одним из важных последствий этих изменений служит
сдвиг мембранного потенциала фоторецепторной клетки.
. Рецепторные потенциалы
Сдвиг мембранного потенциала фоторецепторной клетки (П или К), который
принято называть рецепторным потенциалом, происходит на ее плазмолемме, тогда,
как фотоизомеризация родопсина совершается преимущественно во внутриклеточных
мембранах, то есть, в мембранных дисках наружных сегментов.
Свойство фоторецепторных мембран изменяется одновременно с
конформационной перестройкой в них опсина. После поглощения (кванта света), спустя 1,5-2 мс,
наблюдается гиперполяризация плазмолеммы фоторецепторной клетки (П или К).
Следовательно, в палочке (К) существует механизм передачи сигнала с
фоторецепторных мембран дисков на плазмолемму П (К). В передаче участвует, так
называемый, внутриклеточный посредник, природа которого до настоящего времени
полностью не установлена, однако, считают, что в посредничестве участвуют
циклический гуанозинмонофосфат (цГМФ) и ионы .
В следствие конформационной перестройки молекул родопсина, в
фоторецепторной мембране изменяется содержание внутриклеточного посредника в
цитозоле наружного сегмента П или К. Это приводит к закрытию 100-300 натриевых
каналов в плазмолемме, которые в отсутствие светового раздражителя пребывали в
открытом состоянии. Они обладают свойствами потенциалнезависимых ионных каналов
и, будучи управляемые светом, получили название фотозависимых натриевых
каналов. Когда закрываются 100-300 каналов, электропроводимость плазмолеммы
наружного сегмента падает на 1-3%. Падение мембранной электропроводимости
приводит в гиперполяризации фоторецепторной клетки. Гиперполяризационный сдвиг
мембранного потенциала П (К), возникающий под действием света, называют
рецепторным потенциалом (РП). Принято считать, что ПП фоторецепторной клетки
генерируется в темноте на плазмолемме внутреннего сегмента. По своей природе ПП
является калиевым потенциалом, подобно всякому мембранному потенциалу в
условиях покоя. В темноте ПП палочки (колбочки) составляет примерно от -20 до
-40 мВ.
В системе генерации энергетического потенциала П плазмолемма наружного
сегмента выполняет роль шунта с переменной электропроводностью.
Электропроводность регулируется (управляется) светом при помощи фотозависимых
натриевых каналов. Когда все натриевые каналы плазматической мембраны наружного
сегмента одной П оказываются заблокированными светом, удельное поверхностное
сопротивление этой мембраны достигает величины 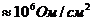 . В темноте ионный ток, входящий
через каналы в цитоплазму, довольно значителен, и составляет примерно 40 nА. А
при освещении сетчатки он блокируется, при этом степень его ослабления зависит
от интенсивности света. Чем слабее натриевый ток через плазматическую мембрану
наружного сегмента, тем больше выражена гиперполяризация в фоторецепторной
клетке. Изменение потенциала П (К) во времени имеет вид:
. В темноте ионный ток, входящий
через каналы в цитоплазму, довольно значителен, и составляет примерно 40 nА. А
при освещении сетчатки он блокируется, при этом степень его ослабления зависит
от интенсивности света. Чем слабее натриевый ток через плазматическую мембрану
наружного сегмента, тем больше выражена гиперполяризация в фоторецепторной
клетке. Изменение потенциала П (К) во времени имеет вид:
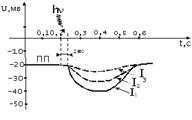
Рецепторные потенциалы одиночной колбочки, возникающие в ответ на
короткие (10 мс) вспышки света при трех разных интенсивностях света
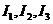 - интенсивность света. Чем больше I, тем больше
гиперполяризация
- интенсивность света. Чем больше I, тем больше
гиперполяризация 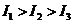 .
.
При поглощении 1 фотона () гиперполяризация фоторецепторов разных представителей
животного мира неодинакова. Так, в палочках сетчатки позвоночных животных
гиперполяризация составляет десятки и сотни мкВ, тогда, как в фоторецепторных
клетках членистоногих она достигает 10 мВ. Эти приведенные числа подтверждают
положение об усилении сигнала в рецепторных аппаратах. Действительно, квант
света, несущий энергию 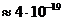 Дж, вызывает фотолиз только одной молекулы родопсина, что в
свою очередь приводит к закрытию в плазмолемме фоторецепторной клетки
позвоночных животных 100-300 натриевых каналов. Это приводит к образованию РП.
Однако, энергия РП на 3 порядка превосходит энергию вызвавшего его фотона.
Дж, вызывает фотолиз только одной молекулы родопсина, что в
свою очередь приводит к закрытию в плазмолемме фоторецепторной клетки
позвоночных животных 100-300 натриевых каналов. Это приводит к образованию РП.
Однако, энергия РП на 3 порядка превосходит энергию вызвавшего его фотона.
Амплитуда рецепторного потенциала возрастает при повышении интенсивности
света, падающего на сетчатку, причем, в пределах 1000-кратного изменения
интенсивности, эта зависимость имеет логарифмический характер:
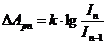 - изменение амплитуды РП.
- изменение амплитуды РП.
- коэффициент пропорциональности;
 - интенсивность последующего;
- интенсивность последующего;
- интенсивность предыдущего.
При ярком освещении амплитуда рецепторного потенциала может достигать 25
мВ. Величина РП зависит также и от длины волны света. П или К в ответ на
зеленый свет (=500 нм), генерируют наибольший РП. Среди К выделено 3 типа:
1) дает максимальный ответ РП на синий
цвет;
2) дает максимальный ответ РП на зеленый
цвет;
3) дает максимальный ответ РП на желто-красный
цвет.
Использование микроэлектронной техники для изучения реакции
фоторецепторов на облучение светом, позволило обнаружить деполяризацию П и К,
при этом деполяризация предшествует гиперполяризации. Амплитуда деполяризации
очень мала и составляет примерно 0,1 мкВ. Учитывая временные соотношения де- и
гиперполяризации, первую из них стали называть ранним, а вторую - поздним РП, и
обозначили, соответственно, РРП и ПРП. Установлено, что РРП обусловлен
преимущественно перемещениями электрических диполей в мембранах наружного
сегмента. Эти перемещения диполей связаны с фотолизом молекул родопсина.
Другими словами, РРП обусловлен током смещения в фоторецепторных мембранах, и
РРП не служит промежуточным звеном передачи сигналов от них к синоптической зоне
П или К, при действии света. Эта роль принадлежит гиперполяризации. В отличие
от других рецепторных аппаратов, где таким звеном служит деполяризационные
сдвиги мембранного потенциала. Таким образом, только поздний РП является
рецепторным потенциалом П или К в полном значении этого термина.
. Распределение сигнала по сетчатке
Поздний РП, возникнув в НС, распределяется в синоптической зоне П или К
по закону электротонической передачи (с декрементом). При этом, постоянная
длины плазмолеммы фоторецепторной клетки, которая проводит гиперполяризацию,
достигает 10 мкм. Достигнув синоптической зоны П (К) ПРП изменяет свойства ее
мембраны и, тем самым, влияет на медиаторную систему, то есть, на передачу
сигнала к биполярному нейрону. Химическая природа медиатора, выделяемого
фоторецепторной клеткой, пока еще неизвестна точно, однако, наиболее вероятными
кандидатами на эту роль признаются глутамат и аспарат. Обе аминокислоты
обладают деполяризующим действием на постсинаптическую мембрану. При облучении
П (К) светом, биполярные нейроны так же испытывают гипреполяризацию.
Принято считать, что фоторецепторной клетке в темноте свойственен
определенный темп выделения деполяризующего медиатора (глутамата или аспарата),
а гиперполяризация синаптической зоны при освещении замедляет темп выброса
медиатора, следовательно, постсиноптические мембраны, принадлежащие биполярным
нейронам, также гиперполяризуются.
Гиперполяризационный сдвиг мембранного потенциала биполярного нейрона
электротонически распространяется по эго-плазмолемме к синапсу на теле третьего
нейрона, который входит в состав афферентного пути сетчатки.
Медиатор, поступающий от биполярного нейрона ганглиозной клетки, вызывает
ее деполяризацию.
Деполяризационный сдвиг потенциала распределяется электротонически с тела
по аксонам ганглиозной клетки. Плазматическая мембрана аксона ганглиозной
клетки (аксолемма) обладает возбудимостью, благодаря чему там и формируется ПД,
когда деполяризация достигает порогового уровня. ПД, неся информацию от
фоторецепторов, поступает в ЦНС по волокнам зрительного нерва. Эти волокна
представляют собой аксоны ганглиозных клеток сетчатки.
Схематически передачу поздних РП по сетчатке можно представить в виде:
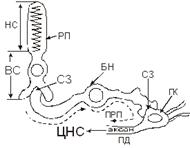
Схема передачи фоторецепторного сигнала по афферентному пути в пределах
сетчатки
РП - рецепторная палочка; НС - наружный сегмент; ВС - внутренний сегмент;
СЗ - синоптическая зона; ГК - ганглиозная клетка; ПРП - поздний рецепторный
потенциал; ПД - потенциал действия; ЦНС - центральная нервная система
Зрительный нерв включает примерно 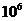 волокон. По этим волокнам в мозг
направляются сигналы от 130
волокон. По этим волокнам в мозг
направляются сигналы от 130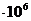 фоторецепторов. Следовательно, в среднем 1 аксон ГК собирает
информацию от множества рецепторных клеток. Все фоторецепторы, посылающие
сигналы к одной ГК, составляют ее рецепторное поле. Рецепторные поля,
образованные П, гораздо многочисленнее, по сравнению с К рецепторными полями.
Необходимо отметить, что каждая К, находящаяся в центральной ямке желтого пятна
сетчатки, связана только с одной ГК. Другими словами, рецепторное поле нейрона
включает только одну колбочку. С этим связана наилучшая разрешающая способность
центрального К зрения. Другие ганглиозные клетки получают сигналы больше, чем
от 1 фоторецептора.
фоторецепторов. Следовательно, в среднем 1 аксон ГК собирает
информацию от множества рецепторных клеток. Все фоторецепторы, посылающие
сигналы к одной ГК, составляют ее рецепторное поле. Рецепторные поля,
образованные П, гораздо многочисленнее, по сравнению с К рецепторными полями.
Необходимо отметить, что каждая К, находящаяся в центральной ямке желтого пятна
сетчатки, связана только с одной ГК. Другими словами, рецепторное поле нейрона
включает только одну колбочку. С этим связана наилучшая разрешающая способность
центрального К зрения. Другие ганглиозные клетки получают сигналы больше, чем
от 1 фоторецептора.
Существуют такие ГК, которые собирают сигнализацию от множества тысяч
палочек, следовательно, рецепторные поля сетчатки весьма разнообразны.
68. Основы фотометрии
Свойства ФР и всей зрительной системы человека оказали весьма большое
влияние на фотометрию. Фотометрия - это раздел физической оптики, в котором
рассматриваются энергетические характеристики светового излучения в процессах
его испускания, распределения и взаимодействия с веществом. К области
фотометрии относится только та часть электромагнитного спектра, которая
воспринимается человеческим глазом. В необъятном спектре электромагнитных
колебаний, только узкая полоса видимой области электромагнитных колебаний с
длиной волн от 380 нм до 760 нм, воспринимается зрительной системой
человеческого глаза, то есть, как принято говорить, обладает видимостью.
Однако, к волнам разной длины, принадлежащим к видимой области,
электромагнитная колебательная чувствительность зрительного анализатора
человека неодинакова, следовательно, различаемые участки видимой области спектра,
обладают разной видимостью. Кривые видности были построены в 20-е годы ХХ века
по результатам определения чувствительности зрительного анализатора человека к
свету разных длин волн. Кривые видности имеют вид:
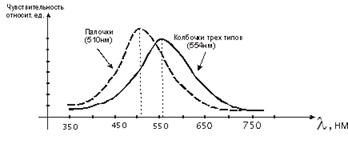
Кривые видности фотопического (сплошная линия) и скотопического
(штриховая линия) зрения
Одна из кривых построена для скотопического (палочкового) зрения
(штрихованная линия). Она почти полностью совпадает со спектром поглощения
родопсина в видимой области спектра. Максимальная чувствительность зрительного
восприятия, когда она обеспечивается П при малой освещенности примерно 0,01
люкса (Лк) приходится на =510 нм (зеленый свет). Другая кривая видимости отображает
спектральную чувствительность фотопического (колбочкового) зрения. Фотопическая
кривая является результатом интегрирования функции спектральной
чувствительности К трех типов. Интегрирование осуществляется в центральном
отделе зрительного анализатора. Максимальная интегральная световая
чувствительность К зрительной системы приходится на =554 нм.
Чувствительность ФР необычно высока, так, возбуждение П (К) происходит
при поглощении его 1 кванта света. Абсолютная световая чувствительность зрительной
системы человека определяется по порогу ощущения ниже чувствительности
отдельного рецептора. Человек ощущает только такой слабый свет, который
одновременно возбуждает не менее 5-10 палочек, для чего им необходимо поглотить
столько же фотонов. В ходе биологической эволюции, П сетчатки достигли предела
световой чувствительности, который допускается квантовой теорией. Этим
обусловлена высочайшая чувствительность к свету всей зрительной системы
человека. Так, пороговая яркость для стахопического зрения составляет 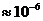 кд/.
кд/.
. Сопоставление энергетических и светотехнических параметров света
Кривая видности фотопического зрения позволяет осуществить приведение
физических величин, характеризующих световую энергию, к, так называемым,
светотехническим величинам, которые выражают ее оценку с учетом особенностей
зрительного восприятия человека.
Две группы названных параметром сведем в таблицу:
|
N
|
Энергетические параметры
света
|
N
|
Психологические параметры
|
|
1
|
Поток излучения (лучистый
поток) 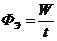 [Вт] [Вт]
|
1
|
Световой поток 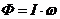 , , 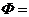 [Люммен] [Люммен]
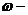 телесный угол в который распространяется свет. телесный угол в который распространяется свет. 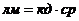 , ср- стерадиан. , ср- стерадиан.
|
|
2
|
Энергетическая сила света 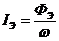 [Вт/ср] - сила излучения. [Вт/ср] - сила излучения.
|
2
|
Сила света I [кд]
|
|
3
|
Энергетическая яркость 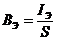 [Вт/ср [Вт/ср ] ]
|
3
|
Яркость 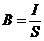 [кд/] [кд/]
|
|
4
|
Энергетическая освещенность
(облученность) 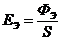 [Вт/] [Вт/]
|
4
|
Освещенность 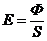 [люкс (лк)]=[лм/] [люкс (лк)]=[лм/]
|
Основным техническим параметром света служит поток излучения - это
энергия квантов света, излучаемая в единицу времени.
Энергетическая сила света представляет собой поток излучения,
приходящийся на единицу телесного угла в данном направлении.
Под энергетической яркостью понимают энергетическую силу света,
отнесенную к единице площади проекции поверхности излучающего тела на
направление, перпендикулярное распределению света.
Энергетическая освещенность - это поток излучения, падающий на единицу
площади облучаемой поверхности. Следовательно, энергетическая яркость является
характеристикой источника излучения, а энергетическая освещенность
характеризует облучаемую поверхность.
За основную светотехническую величину принята сила света. Ее размерность
- [кд] Значение [кд] принимают таким, что яркость полного излучения при
температуре затвердевания Pt (2042 К или  С) равна 60 kд/
С) равна 60 kд/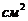 . Применявшаяся ранее международная
свеча составляет 1,005 kд. Производными параметрами от силы света являются
яркость и световой поток. Единицей яркости служит kд/, то есть, одна kд/ характеризует яркость источника
каждого излучающей поверхности, которая
имеет в данном направлении силу света = 1 kд. В медицине широко используются и
внесистемные единицы яркости света: нит, стильб (сб), апостильб (апб), ламберд
(Лб).
. Применявшаяся ранее международная
свеча составляет 1,005 kд. Производными параметрами от силы света являются
яркость и световой поток. Единицей яркости служит kд/, то есть, одна kд/ характеризует яркость источника
каждого излучающей поверхности, которая
имеет в данном направлении силу света = 1 kд. В медицине широко используются и
внесистемные единицы яркости света: нит, стильб (сб), апостильб (апб), ламберд
(Лб).
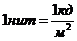 ;
; 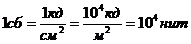 ;
; 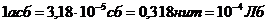
Яркость снега в солнечный день достигает примерно 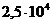 kд/, а яркость земляного грунта
беззвездной ночью - примерно kд/.
kд/, а яркость земляного грунта
беззвездной ночью - примерно kд/.
Световой поток определяется произведением силы света на телесный угол, в
который он распределяется.
Производной величиной от светового потока является освещенность,
представляющая собой световой поток, приходящийся на единицу облучаемой
поверхности. Соотношение между яркостью объекта и освещенностью выражается
формулой:
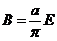
- коэффициент отражения света;
В - яркость;- освещенность.
70. Приведение энергетических параметров к светотехническим
Приведение осуществляется посредством величины, которая получила название
видности. Абсолютной видностью - v - называют соотношение светового потока к
тому энергетическому потоку излучения, который вызывает ощущение данного
потока.
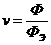
Размерность [v]=[лм/Вт], и отражает субъективную "стоимость" в
лм зрительного ощущения, вызываемого потока излучения в 1 Вт. Известно, что 1
Вт излучения разных вызывает неодинаковые по интенсивности ощущения света. Самым
ярким ощущается свет с = 554 нм. На эту приходится максимум кривой видности
фотопического зрения. Поток излучения в 1 Вт, при = 554 нм, создает у человека
ощущение светового потока в 683 лм, следовательно, максимальная абсолютная видность
равна 683 лм/Вт.
Для излучения на другую , абсолютная видность ниже, что наглядно выражается кривой
видностью. Однако, при решении фотометрических задач, удобно использовать
относительную видность ( ), под которой понимают отношение абсолютной видности при
данной к максимуму видности.
), под которой понимают отношение абсолютной видности при
данной к максимуму видности.
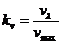
Соответственно, что относительная видность равна 1 при =554 нм. При всех других , относительная видность меньше 1.
71. Биофизические основы вкуса. Восприятия вкуса
В эволюции животных огромное значение имело контактное дифференцированное
распознавание пищи, в результате которой происходило или поедание пищи или
отклонение ее.
Химические соединения, входящие в состав пищи животных, действуя на
вкусовые рецепторы, обусловливают ее соленый, кислый, сладкий или горький вкус.
Имеются более сложные системы квалификации вкусовых ощущений, однако, для
человека предполагают справедливым предположение о наличии только четырех
элементарных вкусовых ощущений.
У человека вкусовые луковицы, содержащие вкусовые рецепторы, находятся во
вкусовых сосочках, которые расположены на поверхности языка, на задней стенке
глотки, в мягком небе, в миндалинах и в надгортаннике. Вкусовые луковицы имеют
овальную форму и содержат, кроме вкусовых клеток, также и опорные клетки.
Вкусовые клетки сообщаются с поверхностью слизистой оболочки с помощью вкусовых
пор.
Рецепторные клетки в опекальной области имеют микровиллы (микроворсинки),
которые в виде кисточки через вкусовые поры сообщаются с жидкой средой
слизистой оболочки. Предполагают, что на поверхности микровилл расположены
активные белковые центры, с которыми реагируют молекулы или ионы вкусовых
веществ. Пространство между микровиллами заполнено особой субстанцией
мукополисахаридной и мукопротеидной природы. Некоторые ученые считают, что эта
субстанция функционирует наподобие ионно-обменной смолы, давая возможность
молекулам или ионам одних веществ контактировать с поверхностью микровилл, а
других - нет. У основания луковицы вкусовые клетки образуют синопсы с
окончаниями афферентных нервных волокон лицевого языкоглоточного и блуждающего
нервов.
В последние десятилетия были проведены большие исследования вкусового
анализатора с помощью микро-ЭВМ. Было установлено, что вкусовые клетки имеют
потенциал покоя ПП величиной, которая изменяется в пределах от -30 до -50 мВ.
При действии вкусового стимула на клетку, наблюдается медленная деполяризация
клеточных мембран, то есть, формируется генераторный потенциал ГП. В
зависимости от интенсивности стимула, его величина обычно меняется в пределах
от 15 до 45 мВ.
Механизм передачи возбуждения от вкусовой клетки к нервному волокну
окончательно не выяснен. Однако, известно, что передача в синапсе
осуществляется при участии ацетилхолина.
Оказалось, что одна и та же клетка может реагировать на сладкие, соленые
и кислые стимулы тогда, как другие клетки обладают повышенной чувствительностью
только к одному или двум стимулам. В отличие от вкусовых клеток, вкусовые
сосочки строго специфичны, при этом, электрическое раздражение отдельных
сосочков вызывает, как правило, одно из четырех вкусовых ощущений. Химическая
стимуляция этих же сосочков у человека вызывает аналогичное ощущение. На основе
подобных опытов, Бекеши в 1966 г. пришел к заключению, что существует полное
совпадение вкусовых качеств для каждого отдельного сосочка, стимулируемого как
электрическим, так и химическим путем. Сосочки отличаются строгой локализацией
на поверхности языка, и делятся на соленые, кислые, сладкие и горькие.
Возможно, что строгая специфичность сосочков обусловлена ионно-обменными
свойствами субстрации, которая заполняет пространство между микровиллами. При
этом, субстрация обусловливает избирательную проницаемость вкусовых веществ.
В опытах с отведением потенциалов от афферентных нервов, было обнаружено,
что небольшое количество нервных волокон обладает узкой вкусовой
специфичностью, а большая часть волокон обладает или относительной
специфичностью или вообще, не обладает специфичностью. Необходимо отметить, что
наличие специфичности в периферическом отделе вкусового анализатора,
обусловливает формирование дифференцированных вкусовых ощущений в центральном
отделе головного мозга. Натуральные пищевые раздражение одновременно возбуждает
несколько или все вкусо-воспринимающие элементы. В результате чего возникает
сложное вкусовое ощущение.
. Химические свойства вкусовых веществ и теория вкуса
Все вкусовые вещества условно можно разделить на три группы:
1. Вещества, близкие по своей химической
структуре и вызывающие примерно одинаковые вкусовые ощущения.
2. Вещества, близкие по химическим
свойствам, но обладающие разным вкусом.
3. Вещества разной структуры, но
сходного вкуса.
К первой группе относятся сахара: глюкоза, сахароза, галактоза, лактоза.
Все они обладают сладким вкусом. Кислым вкусом обладают диссоциированные
неорганические и органические кислоты.
Ко второй группе относится ряд изомеров, например, -валин, имеющий горьковато-сладкий
вкус; 1-валин, имеющий сладкий вкус; d-лейцин, имеющий сладкий вкус; 1-лейцин,
имеющий горький вкус.
К третьей группе можно отнести, например, полисахариды: глицерин,
гликокол, сахарин, нитробензол, хлористый метил, хлороформ, - имеющий сладкий
вкус, а так же, хинин, нитропроизводные бензола и многие неорганические соли и
кислоты, имеющий горький вкус. Аналогичность вкуса при таком несходстве
химической структуры вещества, главным образом, относится к горькому и сладкому
вкусам.
Соленый вкус NaCl присущ и другим хлоридам (K, 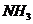 , Ca и т.д.). Можно было бы
предположить, что свободный анион
, Ca и т.д.). Можно было бы
предположить, что свободный анион 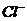 , а не катионы , ,
, а не катионы , , 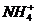 , , ответственен за соленый вкус,
однако, значение катионов также нельзя исключить. Так, например, в слабой
концентрации NaCl обладает соленым вкусом, NaBr - горьковато-соленым, NaI -
солоновато-горьким. Из этого можно заключить, что ион придает растворам всех этих солей
присущий им солоноватый вкус. В настоящее время считают, что в основе соленого
стимула лежит действие низкомолекулярных анионов и катионов. При этом,
принимают, что интенсивность вкуса низкомолекулярных солей зависит от катионов
(, ,
, , ответственен за соленый вкус,
однако, значение катионов также нельзя исключить. Так, например, в слабой
концентрации NaCl обладает соленым вкусом, NaBr - горьковато-соленым, NaI -
солоновато-горьким. Из этого можно заключить, что ион придает растворам всех этих солей
присущий им солоноватый вкус. В настоящее время считают, что в основе соленого
стимула лежит действие низкомолекулярных анионов и катионов. При этом,
принимают, что интенсивность вкуса низкомолекулярных солей зависит от катионов
(, , 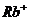 ,
, 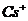 и т.д.), а характер вкуса от анионов
(,
и т.д.), а характер вкуса от анионов
(, 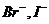 и т.д.).
и т.д.).
Кислым вкусом обладает большинство органических и неорганических кислот.
Как известно, общей чертой всех кислот является их способность к диссоциации в
водных растворах на анионы и катионы . Кислый стимул связывают с
воздействием на вкусовые клетки ионов . В этом убедились при помощи опытов.
Так, например, HCl ощущается кислотой при разбавлении в пропорции 1:800, в то
время, как при этой же концентрации, весь диссоциированный NaCl безвкусен.
Воздействие ионов пропорционально его концентрации, однако, более сильные
кислоты в одинаковой концентрации со слабыми, не всегда вызывают более сильное
вкусовое ощущение. Это объясняется тем, что для ионов проницаемость субстанции окружающей
микровиллы вкусовых клеток имеет низкое значение. Слабые же кислоты в виде
недиссоциированных молекул, достигают микровилл вкусовых клеток, и только затем
диссоциируют. При этом, образовавшиеся ионы взаимодействуют с активными центрами
микровилл.
Сладкий вкус вызывается большим числом органических веществ, которые, как
правило, не диссоциируют. В настоящее время окончательно не выяснено, какие
свойства молекул вызывают сладкий вкус. Многие сладкие вещества являются
многоатомными спиртами с повторяющейся группой 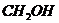 (сахара, глицерин). Сахарин, который
обладает сладким вкусом, имеет совершенно другую структуру. По мнению Акри,
сладкий стимул молекул связан с их возможностью образовывать слабые водородные
связи, с активным центром рецепторной молекулы, которая локализована во
вкусовой клетке. Во всех случаях, структура молекулы должна соответствовать
стерическому положению, и возможностям водородных связей рецепторной молекулы.
(сахара, глицерин). Сахарин, который
обладает сладким вкусом, имеет совершенно другую структуру. По мнению Акри,
сладкий стимул молекул связан с их возможностью образовывать слабые водородные
связи, с активным центром рецепторной молекулы, которая локализована во
вкусовой клетке. Во всех случаях, структура молекулы должна соответствовать
стерическому положению, и возможностям водородных связей рецепторной молекулы.
Менее всего изучена природа горького стимула вкусовых веществ. Было
отмечено, что соли, по мере увеличения молекулярного веса, становятся все более
горькими. Так, например, раствор NaCl имеет соленый вкус, цезий-хлор - горький,
KI - горький. Имеются данные, что горький стимул связан с возможностью
образования слабых дисперсионных связей типа водородных.
Существует несколько теорий, которые пытаются объяснить конкретные
процессы взаимодействия молекул вкусовых веществ с рецепторными клетками. Так,
Лазарев еще в 1920 г., исходя из роли ионов в процессе возбуждения, допускал,
что вкусовые луковицы каждого сосочка содержат высокочувствительные вещества
белковой природы, которые разлагаются под влиянием адекватного стимула. При
этом ионизируемые продукты распада возбуждают соответствующие нервные
окончания.
В настоящее время наиболее разработанной является теория Бейдлера. Он
предположил, что вкусовые стимулы (молекулы или ионы) взаимодействуют с
определенными участками плазматической мембраны вкусовой клетки, которые он
выделяет в качестве активных центров. Активные центры представляют собой
полиэлектролиты белковой природы, содержащие большое количество заряженных боковых
цепей. Взаимодействие вкусовых стимулов с активными центрами протекает по типу
мономолекулярной реакции, которая имеет вид:
С+(N-Z)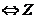 (1)
(1)
С - концентрация стимулирующего вкусового вещества;- общее число активных
центров рецепторов;- число связанных активных центров, при концентрации
вкусового вещества, равного С.
Согласно закона действующих масс, постоянная равновесия данной реакции:
=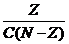 (2)
(2)
Величина ответа рецепторной клетки прямо пропорциональна числу активных
связанных центров R=Z, а максимальный отклик имеет место в том случае, когда
все центры заняты. Тогда уравнение (2) можно записать в виде:
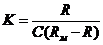
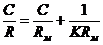 (3)
(3)
Уравнение (3) представляет собой основное уравнение вкусовой рецепции.
Величина C/R выражает суммарную реакцию вкусовых рецепторов. Согласно данному
уравнению, между C/R и С существует линейная зависимость. Данное положение
получило экспериментальное подтверждение.
Бейдлер предполагал, что взаимодействие частиц вкусового вещества с
активными центрами рецептора, представляет собой физический процесс адсорбции,
в основе которого лежит слабое дисперсионное взаимодействие. Это предположение
согласуется с предсказанием Акри об образовании водородных связей между
молекулами сладкого стимула и рецептора. На основании экспериментальных данных
и уравнения вкусовой рецепции (3), можно найти величину (константы) равновесия
конкретной реакции. Она изменяется от 7,7 для бутирата натрия до 9,8 для NaCl.
Эти величины - константы равновесия, очень близки к коэффициенту для адсорбции
солей на белках. Это, в свою очередь, соответствует исходному допущению
Бейдлера об адсорбции вкусового вещества на поверхности рецептора.
О физической природе процесса свидетельствует также и то, что реакция
вкусовых рецепторов на соленый стимул не изменяется при повышении температуры
от 20 до  С. Кроме того, величина реакции
заметно не изменяется при сдвиге pH от 3 до 11, что исключает возможность
сильного электростатического взаимодействия с иногенными группами белковых
молекул. Термодинамические расчеты показывают, что процесс адсорбции частиц
приводит либо к уменьшению количества гидратационной воды на поверхности
рецептора, либо к небольшому изменению конформации молекул рецептора. Возможно,
что изменение конформаций рецепторных молекул лежит в основе дальнейшего
процесса, приводящего в конечном счете к возбуждению вкусовой клетки.
С. Кроме того, величина реакции
заметно не изменяется при сдвиге pH от 3 до 11, что исключает возможность
сильного электростатического взаимодействия с иногенными группами белковых
молекул. Термодинамические расчеты показывают, что процесс адсорбции частиц
приводит либо к уменьшению количества гидратационной воды на поверхности
рецептора, либо к небольшому изменению конформации молекул рецептора. Возможно,
что изменение конформаций рецепторных молекул лежит в основе дальнейшего
процесса, приводящего в конечном счете к возбуждению вкусовой клетки.
Дальнейшее развитие теории вкусового восприятия было связано с открытием
Дастоли сладкочувствительных и горькочувствительных белков во вкусовых
луковицах млекопитающих. Дастоли удалось выделить из специфических
сладкочувствительных и горькочувствительных сосочков белки, которые
избирательно взаимодействуют со сладкими или горькими веществами.
Термодинамическая и кинетическая особенности взаимодействия сладких и горьких
веществ с молекулами выделенных белков, совпадают с особенностями
мономолекулярных реакций, описанных Бейдлером. В этой связи в настоящее время
считают, что именно этими белками представлены активные центры вкусовых клеток,
однако, локализация этих белков во вкусовых клетках еще не установлена.
73. Биофизические основы восприятия запаха
В основе обоняния лежит взаимодействие молекул пахучих веществ с
обонятельными клетками. Молекулы пахучих веществ, отделяясь от своей основной
массы и передвигаясь потоками воздуха, могут действовать на расстоянии.
Обонятельные рецепторы входят в состав обонятельного эпителия,
расположенного в задней части носовой перегородки и в верхнем носовом ходе.
Рецепторные клетки представляют собой биполярные нейроны d примерно 5-10 мкм,
расположенные вокруг цилиндрических опорных клеток. У человека количество
обонятельных рецепторов (ОР) достигает примерно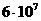 , а площадь обонятельного эпителия
(ОЭ) составляет примерно 5. Поверхность ОЭ покрыта водянистой средой. Периферические
отростки обонятельных клеток заканчиваются на своей вершине утолщение
грушевидной формы, которое называют обонятельной булавой. Как показали
электронно-микроскопические исследования, на поверхности каждой обонятельной
булавы (ОБ) расположено от 9 до 16 очень тонких ресничек. Центральные отростки
отходят от противоположного конца обонятельных клеток и образуют тонкие нервные
нити, которые вступают в полость черепа. На нижней поверхности лобной доли
нервные нити сходятся, образуя обонятельную луковицу, в клубочках которой и
заканчивается первый нейрон обонятельного афферентного пути.
, а площадь обонятельного эпителия
(ОЭ) составляет примерно 5. Поверхность ОЭ покрыта водянистой средой. Периферические
отростки обонятельных клеток заканчиваются на своей вершине утолщение
грушевидной формы, которое называют обонятельной булавой. Как показали
электронно-микроскопические исследования, на поверхности каждой обонятельной
булавы (ОБ) расположено от 9 до 16 очень тонких ресничек. Центральные отростки
отходят от противоположного конца обонятельных клеток и образуют тонкие нервные
нити, которые вступают в полость черепа. На нижней поверхности лобной доли
нервные нити сходятся, образуя обонятельную луковицу, в клубочках которой и
заканчивается первый нейрон обонятельного афферентного пути.
При действии обонятельных стимулов на обонятельные рецепторы, происходит
формирование генераторных потенциалов, которые вызывают в центральных отростках
рецепторов нервные импульсы. Суммарный генераторный потенциал ОР, то есть,
электроольфактограмму, можно зарегистрировать с помощью электрода, приложенного
к поверхности ОЭ. Вопрос ОБ избирательности ОР к молекулам пахучих веществ
окончательно не выяснен. До настоящего времени не было обнаружено строгой
избирательности рецепторов. Так, например, некоторыми учеными с помощью
микроэлектродов регистрировались потенциалы одиночных волокон обонятельного
нерва лягушки, при действии различных пахучих веществ. Все исследуемые
рецепторные нейроны обнаружили грубую избирательную чувствительность запаха.
При этом, каждый из них, отвечал на одни запахи, не отвечал на другие.
Большинство из них дает особо сильный ответ, по крайней мере, на 1 из 25
использованных запахов, и более слабо отвечает на остальные.
. Теория обоняния
В основе возникновения обонятельного ощущения лежит взаимодействие
молекул или частиц пахучего вещества с ОР, однако, до настоящего времени
неясно, какие свойства молекул обусловливают это взаимодействие. Не ясно также,
с молекулами каких веществ, локализованных в рецепторных клетках, происходит
взаимодействие. Попытки объяснить пахучие свойства молекул их химическими
свойствами и структурой, не увенчались успехом. Вещества со сходными свойствами
и структурой, могут обладать разными запахами и наоборот. В настоящее время, из
всех теорий восприятия запаха, наибольшее внимание заслуживают две:
1) квантовая;
2) стереохимическая.
Согласно квантовой теории запаха, молекулам пахучего вещества свойственны
внутримолекулярного колебания, в результате которых они испускают
электромагнитное излучение в инфракрасной области спектра (ИК). Это излучение
взаимодействует с молекулами обонятельного рецептора пигмента, типа
каротиноидов, находящихся в мембране рецепторов. По мнению авторов данной
теории, молекулы веществ с похожими запахами, должны характеризоваться сходными
НЧ колебаниями. В качестве подтверждения, Райд приводит вещества, обладающие
миндальным запахом и близкой частотой колебаний: нитробензол, бензонитрил, бутиронитрил
и т.д. из обонятельных клеток удалось выделить ряд каротиноидов и витамин А,
которые, по мнению Райда, поглощают электромагнитные излучения молекул пахучих
веществ.
Однако, несмотря на известные успехи, данная теория встречает ряд очень
серьезных возражений. Так, многие вещества со сходными запахами имеют разные
частоты колебаний и разные спектры поглощения в ИК области (например, многие
спирты). Молекулы, в которых атом водорода замещен изотопом дейтерия, обладают
сходными запахами. Хотя, при этом значительно меняются частоты основных
колебаний, с другой стороны молекулы с почти одинаковыми частотами колебаний и
спектрами ИК поглощения, обладают несходными запахами, кроме того, не
установлена локализация пигментов в обонятельных клетках и не доказано их
участие в первичном процессе взаимодействия с пахучими молекулами.
Более обоснованной является стереохимическая теория восприятия запаха,
выдвинутая Монкрифом и подробно разработанная Эймуром. Согласно этой теории,
запах вещества обусловлен не химическим составом молекулы, а их формой и
размерами. Обонятельная система состоит из рецепторов разных типов, каждый из
которых соответствует отдельному первичному запаху. Поверхности рецепторных
клеток имеют углубления (лунки) определенной формы. Молекулы пахучих веществ
вызывают ощущение запаха только в том случае, если их форма соответствует форме
лунок рецепторов, в которые они плотно входят. Этот принцип аналогичен принципу
"замка и ключа", при взаимодействии фермента и субстрата. Согласно представлениям
Эймура, вещества, имеющие сходный запах, должны обладать сходной формой своих
молекул. Эймур, сопоставив большое количество данных о форме молекул с
ощущениями, которые они вызывают, пришел к выводу о наличии 7 первичных простых
запахов:
·
камфорноподобного;
·
мускусного;
·
цветочного;
·
мятного;
·
эфирного;
·
острого;
·
гнилостного.
В этом смысле, первичные запахи аналогичны трем основным цветам
зрительного и четырем вкусам вкусовых анализаторов. Каждому первичному запаху
соответствует определенная форма молекулы и лунки на поверхности рецептора.
Так, эфирная молекула отличается палочковой формой, при этом, она должна иметь
вытянутую лунку с размерами: длина = 1,8 нм, ширина = 0,5 нм, глубина = 0,4 нм.
Камфорная молекула имеет сферическую форму, диаметром 0,7 нм, а ее лунка имеет
форму эллиптической чаши, глубиной 0,4 нм, длиной = 0,2 нм, шириной = 0,75 нм.
Молекулы, вызывающие другие ощущения, имеют более сложную форму. Если сложная
молекула внедряется сразу в две лунки, то возникает сложный запах.
Теория Эймура нашла свое подтверждение, что, в некоторых случаях,
оказывалось возможным предсказывать запах веществ, исходя из формы их молекул.
Так было синтезировано несколько органических веществ, была рассчитана их
вероятная форма и предсказан их запах. Однако есть недостатки: острые и
гнилостные запахи не вмещаются в стереохимическую схему. Для молекул,
вызывающих острые и гнилостные запахи, форма и размер не имеют значения.
Решающую роль играет электрический заряд. Острые едкие запахи свойственны тем
соединениям молекулы, которые из-за нехватки электронов, имеют "+"
заряд и сильное сродство к электронам. Гнилостные запахи, наоборот, вызываются
молекулами, обладающими избытком электронов.
Необходимо заметить, что данная теория обходит молчанием вопрос о том,
что происходит после попадания молекулы в соответствующую лунку на поверхности
рецептора? С чем она взаимодействует в ней при этом? Теория не объясняет, как
возникают потенциалы в ответ на заполнение лунок?
. Электрические и магнитные свойства тканей организма
Современные преставления о магнитных свойствах живых тканей основаны на
фактах о молекулярной организации БМ и, в значительно меньшей степени, на
сведениях о кванотовомеханических свойствах физиологически активных молекул.
Характеризуя электрические свойства живых тканей, следует учитывать, что они
являются композиционными средами, поскольку одни структурные элементы обладают
свойствами проводников, а другие - диэлектриков. Предполагают, что ряд
биологически важных макромолекул проявляет полупроводниковые свойства.
. Электропроводность живых тканей
Электропроводность органов и тканей связана с присутствием в них ионов,
которые являются свободными зарядами, создающими в организме ток проводимости
под действием ЭМП, излучаемыми как внешними источниками, так и генерируемых
живыми клетками. Электропроводность живых тканей определяется прежде всего
электрическими свойствами крови, лимфы, межклеточной жидкости и цитозоля.
Удельная электропроводность () этих электролитов составляет 0,1-1 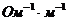 . Подвижность ионов в биологических
жидкостях примерно такая же, как в растворах соответствующих солей,
приготовленных на дистиллированной воде.
. Подвижность ионов в биологических
жидкостях примерно такая же, как в растворах соответствующих солей,
приготовленных на дистиллированной воде.
Однако целых органов на 4-6 ниже жидкостей, выделенных из них.
Причиной не совпадений результатов измерений являются малые объемы, занимаемые
свободными электролитами в органах и тканях животных. В клетке электролиты
заключены в мельчайшие отсеки (компартаменты), образованные БМ, составляющие
более 50% массы клетки. По существу каждая клеточная органелла представляет
собой компартамент. Ее содержимое и окружающий цитозоль обладают относительно
высокой электропроводностью, тогда как разделяющая их мембрана является
типичным диэлектриком.
Удельное сопротивление плазматических мембран измерить достаточно трудно,
так как невозможно измерить толщину мембраны. В этой связи удается измерить
только удельное поверхностное сопротивление. У различных клеток удельное
поверхностное сопротивление изменяется в пределах от 0,4 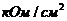 до 30 .
до 30 .
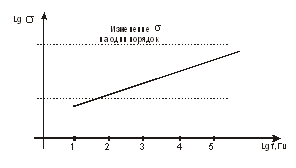
Живым тканям свойственна зависимость электропроводности от частоты
воздействующего ЭМП. Это явление получило название дисперсии
электропроводности. С повышением частоты электропроводность тканей
увеличивается. Дисперсия электропроводности присуща всем средам, а не только биологическим.
Она наблюдается в том диапазоне частот ЭМП, которые соответствуют
характеристическим частотам (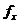 ) заряженных частиц, входящих в состав той или иной среды.
Поскольку однородные среды образованы частицами с близкими значениями , то дисперсия электропроводности в
них выражена слабо. В этой связи сопротивление резисторов в цепях переменного
тока называют активным, в отличие от реактивных сопротивлений емкости и
индуктивности (
) заряженных частиц, входящих в состав той или иной среды.
Поскольку однородные среды образованы частицами с близкими значениями , то дисперсия электропроводности в
них выражена слабо. В этой связи сопротивление резисторов в цепях переменного
тока называют активным, в отличие от реактивных сопротивлений емкости и
индуктивности (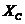 и
и 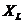 ), которые гораздо сильнее зависят от частоты, чем R. По той
же причине редко утверждается, будто дисперсия электропроводности -
специфическое свойство биологических систем. На самом же деле у них зависимость
), которые гораздо сильнее зависят от частоты, чем R. По той
же причине редко утверждается, будто дисперсия электропроводности -
специфическое свойство биологических систем. На самом же деле у них зависимость
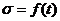 гораздо отчетливее, чем у сред с
менее сложной организацией, и обнаруживается в широком частотном диапазоне. Это
обусловлено сложной мембранной структурой тканей и большим разнообразием
релаксационных способностей их заряженных частиц, причем такое разнообразие
связано, как с различием в размерах, так и с влияние на их подвижность БМ.
Повреждение клеточных мембран стирает в значительной мере грань между тканями и
органическими электролитами в дисперсии на низких частотах.
гораздо отчетливее, чем у сред с
менее сложной организацией, и обнаруживается в широком частотном диапазоне. Это
обусловлено сложной мембранной структурой тканей и большим разнообразием
релаксационных способностей их заряженных частиц, причем такое разнообразие
связано, как с различием в размерах, так и с влияние на их подвижность БМ.
Повреждение клеточных мембран стирает в значительной мере грань между тканями и
органическими электролитами в дисперсии на низких частотах.
. Диэлектрические свойства живых тканей
Диэлектрические свойства биологических тканей определяются присутствием в
них воды, растворенных в ней макромолекул, а также компартамезацией клеточных
структур. Компартамезация, обеспечивая оптимальные условия для внутриклеточных
биохимических процессов, вместе с тем придает живым тканям сегнетоэлектрические
свойства. Показано, что физические среды, в которых чередуются слои очень
низкой и высокой , обладают сегнетоэлектрическими свойствами, если по обе
стороны каждого слоя с низкой присутствуют разные электрические заряды. Такие структуры
подобны доменам с одинаковой ориентацией дипольных моментов. Цитозоль и
содержимое многих органелл обладают относительно высокой 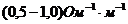 , а БМ - крайне низкой (порядка
, а БМ - крайне низкой (порядка 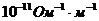 ). Поэтому каждая органелла на БМ
которой поддерживается разность потенциалов между цитозолем и ее содержимым,
имеет значительный дипольный момент подобно домену в сегнетоэлектрике. За счет
таких заряженных компартаментов живые ткани обладают высокой . В постоянном электрическом поле оно
достигает десятка тысяч.
). Поэтому каждая органелла на БМ
которой поддерживается разность потенциалов между цитозолем и ее содержимым,
имеет значительный дипольный момент подобно домену в сегнетоэлектрике. За счет
таких заряженных компартаментов живые ткани обладают высокой . В постоянном электрическом поле оно
достигает десятка тысяч.
Как и всякому домену, каждому внутриклеточному компартаменту присуща не
высокая характеристическая частота релаксации. Применительно к диполям 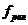 соответствует максимальной частоте
ЭМП, которую они способны воспроизвести своими поворотами в нем, за счет чего
достигается максимально возможная компенсация внешнего поля собственным ЭП
диэлектрика. Диапазон разных внутриклеточных компартаментов находиться в пределах
от Гц до 1 кГц. Поэтому компартаменты вносят основной вклад в диэлектрические
свойства биологических тканей именно на низких частотах.
соответствует максимальной частоте
ЭМП, которую они способны воспроизвести своими поворотами в нем, за счет чего
достигается максимально возможная компенсация внешнего поля собственным ЭП
диэлектрика. Диапазон разных внутриклеточных компартаментов находиться в пределах
от Гц до 1 кГц. Поэтому компартаменты вносят основной вклад в диэлектрические
свойства биологических тканей именно на низких частотах.

На более высоких частотах диэлектрические свойства определяются полярными
макромолекулами, сосредоточенными как в цитоплазме, так и в клеточных
мембранах. В СВЧ полях основной вклад в эти свойства вносит вода.
У разных белковых молекул охватывает диапазон от 10 кГц до 100 мГц и зависит от их
размеров, а также от вязкости окружающей среды. Эту зависимость выражает
формула:
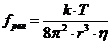 ,
,
где K - постоянная Больцмана; T - температура среды, в которой вращаются
молекулы, под действием ЭМП; - вязкость этой среды; -радиус полярной молекулы.
Важно отметить, что молекулы одинаковых размеров, пребывая в средах с
разной вязкостью (например в цитозоле и БМ) обладают не одинаковой .
Характеристическая частота релаксации внутриклеточный воды такая же, как
и дистиллированной (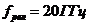 ). Дипольные моменты у них также одинаковы (~1,84 дебая).
Именно в воде, входящей в состав живых тканей, происходят основные
диэлектрические потери при действии на организм СВЧ излучений поскольку
). Дипольные моменты у них также одинаковы (~1,84 дебая).
Именно в воде, входящей в состав живых тканей, происходят основные
диэлектрические потери при действии на организм СВЧ излучений поскольку 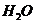 совпадает частота сантиметровых
радиоволн.
совпадает частота сантиметровых
радиоволн.
Неодинаковые величины разных тканевых компартаментов, способных поляризоваться в
ЭМП, обуславливают неравномерный ход кривой  , отображающей зависимость
, отображающей зависимость  ткани от
ткани от 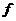 ЭМ колебаний, воздействующих на нее.
На зависимости можно выделить три участка, где кривая идет круче, чем в промежутках.
Эти участки называют зонами дисперсии (релаксации) и их обозначают греческими
буквами
ЭМ колебаний, воздействующих на нее.
На зависимости можно выделить три участка, где кривая идет круче, чем в промежутках.
Эти участки называют зонами дисперсии (релаксации) и их обозначают греческими
буквами 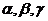 .
.
Первый участок (- дисперсия) соответствует низкочастотному диапазону до 1
кГц. Он обусловлен поляризацией внутриклеточных компартаментов, с которыми
связаны сегнетоэлектрические свойства живых тканей. В силу значительной
инерционности релаксационных процессов в доменах - компартаментах вращение этих
гигантских диполей запаздывает относительно направления 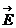 внешнего ЭМП даже на низких
частотах, что проявляется в уменьшении по мере увеличения . Некоторый вклад в - дисперсию вносит релаксация
зарядов на фасциях, внутриорганных соединительно тканных прослойках, клеточных
поверхностях.
внешнего ЭМП даже на низких
частотах, что проявляется в уменьшении по мере увеличения . Некоторый вклад в - дисперсию вносит релаксация
зарядов на фасциях, внутриорганных соединительно тканных прослойках, клеточных
поверхностях.
Второй участок (- дисперсия) отображает изменение поляризации макромолекул по
мере повышения частоты внешнего ЭМП. В скелетной мышце -дисперсия наблюдается в диапазоне
частот от 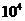 до
до 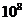 Гц. Снижение по мере повышения в этом диапазоне зависит от того,
что все менее крупные макромолекулы не успевают поворачиваться в соответствии с
частотой внешнего ЭМП, когда она не успевает превосходить
Гц. Снижение по мере повышения в этом диапазоне зависит от того,
что все менее крупные макромолекулы не успевают поворачиваться в соответствии с
частотой внешнего ЭМП, когда она не успевает превосходить  той или иной полярной молекулы.
Очевидно, что Гц является частотой соответствующей наименее инерционных пептидных
молекул.
той или иной полярной молекулы.
Очевидно, что Гц является частотой соответствующей наименее инерционных пептидных
молекул.
Третий участок (- дисперсия) приходиться на частоты выше Гц, чему соответствует . Поскольку воде свойственно
несколько значений лежащих около 20 ГГц (явление многоструктурной воды), то
изменение уменьшиться потому, что даже такие мелкие молекулы, как , не успевают совершать повороты с
частотой, соответствующей частотному диапазону -дисперсии.
На более высоких частотах (до 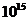 Гц) может проявляться электронная
поляризация, характерная для атомов, и обусловленная смещением оболочек у
атомов.
Гц) может проявляться электронная
поляризация, характерная для атомов, и обусловленная смещением оболочек у
атомов.
В живых тканях, ввиду того, что им наряду с электропроводностью присущи
другие свойства, под действием внешнего ЭМП возникают и токи проводимости, и
токи смещения. По мере повышения частоты ЭМП роль тока смещения в биологическом
эффектах электромагнитного излучения возрастает и становиться ведущей на
частоте выше Гц. Сказанное выше хорошо характеризует различные виды
высокочастотной электротерапии: если при диатермин (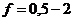 МГц) тепловой эффект связан прежде
всего с током смещения.
МГц) тепловой эффект связан прежде
всего с током смещения.
Клеточные мембраны принято рассматривать по аналогии с диэлектриком в
виде плоских конденсаторов. В этой связи поведение мембран при воздействии ЭМП
характеризует электрическая емкость, приведенная к 1 мембранной поверхности. Эта величина
(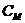 ) называется удельной емкостью БМ и
имеет размерность
) называется удельной емкостью БМ и
имеет размерность 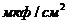 . Очень велика плазмолемы гладкомышечного волокна
млекопитающих - около 30 . Нервные волокна многих животных имеют плазматические
мембраны, … которых находиться в пределах единиц .
. Очень велика плазмолемы гладкомышечного волокна
млекопитающих - около 30 . Нервные волокна многих животных имеют плазматические
мембраны, … которых находиться в пределах единиц .
Чтобы учесть зависимость энергетических преобразований на емкости от
частоты внешнего ЭМП, используется понятие емкостного сопротивления:
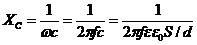

Эта величина служит коэффициентом преобразования энергии внешнего ЭМП в
электрическую энергию, накапливаемую конденсатором в течение половины периода
воздействующих ЭМ колебаний. 4 см больше … БМ, тем меньший электрический заряд
она способна удержать на себе и тем меньшая доля энергии внешнего ЭМП
превратиться в электрическую энергию.
. Магнитные свойства живых тканей
Любая среда является одновременно электриком (проводником или
диэлектриком) и магнитиком. Магнитные свойства биологических тканей
характеризуются довольно низкой величиной магнитной проницаемости ( ) близкой к 1, поскольку основные
химические компоненты биосред (белки, углеводы, липиды, вода) относятся к
диамагнетикам. Их почти нулевая магнитная восприимчивость (
) близкой к 1, поскольку основные
химические компоненты биосред (белки, углеводы, липиды, вода) относятся к
диамагнетикам. Их почти нулевая магнитная восприимчивость ( ) служит одной из причин
недостаточного внимания к изучению магнитных процессов в организме. У человека
обнаружены ферритинсодержащие включения (в надпочечниках). Их функции остаются
не выясненными. Предполагают, что подобные ферромагнитные включения
присутствуют в тканях пчел, бабочек, дельфинов, обеспечивая их пространственную
ориентацию. Вопрос о ферромагнитных свойствах биологических систем далеко от
его разрешения.
) служит одной из причин
недостаточного внимания к изучению магнитных процессов в организме. У человека
обнаружены ферритинсодержащие включения (в надпочечниках). Их функции остаются
не выясненными. Предполагают, что подобные ферромагнитные включения
присутствуют в тканях пчел, бабочек, дельфинов, обеспечивая их пространственную
ориентацию. Вопрос о ферромагнитных свойствах биологических систем далеко от
его разрешения.
Вместе с тем в исследованиях биомагнетизма недостаточно учитывается важное
положение электродинамики, согласно которому коэффициентом преобразования
энергии ЭМП в магнитную энергию среды, подвергнутой его воздействию, является
не сама по себе, а индуктивность,
отображающая как собственно магнитные свойства, так и ее геометрические
особенности. Даже диамагнетики способны осуществлять подобное преобразование
довольно эффективно, если образуют структуры в форме катушек, по которым течет
электрический ток, наведенный внешним полем. (Аналог магнитные поля катушек из
медного провода) Для учета зависимости энергетических преобразований на
индуктивности на частоте внешнего ЭМП введено понятие индуктивного
сопротивления:
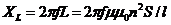
где - длина катушки, по которой течет ток; S - площадь каждого
витка; n - число витков в катушке.
Оказалось, что 1 мембраны аксона кальмара толщиной примерно 10 нм имеет  Гн. Такая индуктивность присуща
медному проводу длиной в 1 милю (1600м), намотанный на железный сердечник
массой в 1 фунт (~450гр). Это очень высокая индуктивность.
Гн. Такая индуктивность присуща
медному проводу длиной в 1 милю (1600м), намотанный на железный сердечник
массой в 1 фунт (~450гр). Это очень высокая индуктивность.
И все же биофизические основы биомагнитных явлений еще не изучены, хотя с
давних пор люди верили в действие магнитных полей на биологические системы. О
лечебных свойствах магнита писал Аристотель (IV в до н.э.) Гален (III в до
н.э.) применял магнит как средство от запоров. Авиценна (XI в) воздействовал на
патологические процессы в селезенке Парацельс (XIV в) применял магнит при
многих болезнях, будучи уверенным в том, что "… магнит оттягивает грыжу и
исцеляет перелом, вытягивает желтуху и оттягивает водянку".
С.П. Боткин в 70 годах прошлого века утверждал, что магнит может
создавать ощущение зуда, покалывания или боли, восстанавливать нарушенную
чувствительность кожи, купировать судороги, вызывать общую слабость и
сонливость. В зависимости от исходного состояния пациента магнит зачастую
приводил к эффектам, противоположным тем, на которые рассчитывал врач, например
вместо ослабления боли усиливал ее.
Современная медицинская литература богата сообщениями о лечебном действии
магнитного поля при атеросклерозе и гипертонической болезни, бронхиальной
астме, неврозе и многих других патологических состояниях. Однако нельзя быть
уверенным в том, что в благоприятном действии магнита нет больше доли
психотерапевтического воздействия на больного самой процедурой магнитотерапии.
До сих пор в биофизике нет достаточно обоснованной рабочей гипотезы, которую
можно положить в основу научного изучения биомагнетизма.
. Дисперсия импеданса живых тканей
Отражением современного состояния магнитобиологии является общепринятое
мнение, будто электрический импеданс (полное сопротивление переменому току)
живых тканей имеет только две состаляющие - резистивное и емкостное
сопротивление:
Индуктивное сопротивление не учитывается.
Как и в электрических цепях, импеданс биологических систем зависит от
частоты переменного тока. Для живых тканей характерно уменьшение импеданса по
мере увеличения этой частоты. Зависимость Z от получила название дисперсии
импеданса. Между дисперсиями электропроводности, диэлектрической проницаемости,
о которых идет речь шла выше, и дисперсией импеданса существует связь, но это
не идентичные процессы. Например, крутые и пологий участки дисперсий и Z в одной и той же ткани обычно не
совпадают. Дисперсия импеданса отображает, очевидно, более широкий круг
электромагнитных процессов в биологических системах (возможно и индуктивные
свойства). Во всяком случае, она сильнее зависит от разнообразных нарушений
жизнедеятельности тканей.
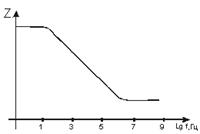
По кривой дисперсии импеданса возможно судить об уровне обмена веществ и
его отклонениях от нормы. В медико-биологических экспериментах и клинике все
шире применяется метод изучения дисперсии Z для оценки жизнеспособности органов
и тканей. Обычно измеряют всего два значения Z: на низкой (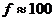 Гц) и высокой (
Гц) и высокой (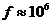 Гц) частотах, соответствующим тем
частотным диапазонам, где кривая дисперсии импеданса идет более полого, чем на
среднечастотном участке крутого спада. Отношение этих величин называют
коэффициентом поляризации (
Гц) частотах, соответствующим тем
частотным диапазонам, где кривая дисперсии импеданса идет более полого, чем на
среднечастотном участке крутого спада. Отношение этих величин называют
коэффициентом поляризации (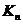 ):
):
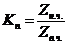
где 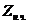 - импеданс на низкой частоте;
- импеданс на низкой частоте; 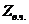 - на высокой частоте.
- на высокой частоте.
Жизнеспособная ткань имеет 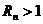 , причем тем больше, чем выше уровень обмена
веществ в данной ткани и чем лучше сохраняется ее структурная целостность. При
отмирании ткани ее стремиться к 1.
, причем тем больше, чем выше уровень обмена
веществ в данной ткани и чем лучше сохраняется ее структурная целостность. При
отмирании ткани ее стремиться к 1.
Кроме используют, так называемый коэффициент частотной дисперсии (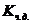 ):
):
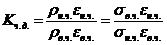
Метод исследования дисперсии импеданса применяют для оценки
жизнеспособности тканевых трансплантантов при пересадке органов. Изучают
возможности использования его для определения зон раневого процесса в ходе
хирургической обработки раны, для характеристики ишемии, отека и т.д.
Широкое распространение в медицинской практике нашла методика
реоплетизмографии. Посредством ее изучают активную составляющую импеданса (R),
которая зависит прежде всего от кровенаполнения исследуемого органа. Чем больше
крови содержится в органе, тем ниже (при прочих равных условиях) его
электрическое сопротивление. Это позволяет оценивать органное кровообращение
путем измерения R органа переменному току.
По динамике электрического сопротивления кожи судят о так называемых
кожно-гальванических реакциях (КГР), в которых отображаются эмоции, утомление и
другие состояния организма. Наконец специалисты в области рефлексотерапии
измеряют электрическое сопротивление кожи для нахождения "активных
точек", воздействие на которые оказывают разнообразные лечебные эффекты.
Есть и другие применения в медицинской практике тех или иных способов измерения
электрического импеданса, его составляющих и арсенал различных методов
исследования электрических свойств биологических систем неуклонно нарастает.